|
Třída
Trilobita - trilobiti |
Charakteristika
Trilobiti (Trilobita: název odvozen z
latinského tres
= tři, lobus = lalok) jsou vyhynulou
skupinou dorzoventrálně zploštělých
členovců. V dospělosti dosahoval velikosti
|
|
Obr. 1: Morfologické znaky hřbetního
krunýře trilobitů (podle Clarksona, |
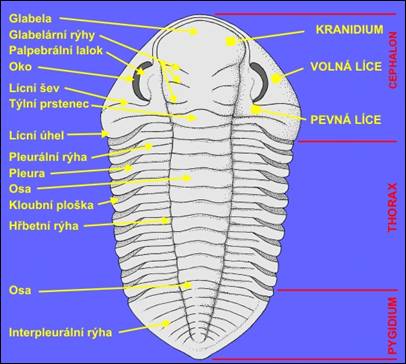
Hlavový
štít bývá půlkruhovitého až
trojúhelníkovitého tvaru. Osní část se
označuje jako glabela (bývá
výrazně vypuklá), která bývá
členěna glabelárními rýhami do glabelárních laloků (obr. 2). Jejich
uspořádaní a počet odpovídá
původní segmentaci cephalonu. Rýhy mohou být
průběžné, nebo kratší, nesouvislé rýhy laterální.
Glabela může být po celém povrchu hladká nebo
zrnitá. Na bázi glabely se nachází tylní
prstenec. Před glabelou může být vyvinuto preglabelární pole. To je
rozděleno na okrajový lem nebo val a líce. Líce lze obvykle dělit a líce vnitřní
(pevné) a líce
vnější (hybné). Rozhraní mezi nimi
vytvářejí lícní švy. Pevné
líce se přimykají ke glabele a společně
s preglabelárním polem vytvářejí celek
označovaný jako kranidium.
Průběh lícních švů je v zásadě
trojího typu. Rozeznává se propární (zadní část
protíná okraj cephalonu před lícním
úhlem nebo trnem), opistopární
(protíná zadní okraj kranidia) nebo běží
po hraně kranidia (marginální šev). Na
párových, méně klenutých bočních
lalocích kranidia (pevné líce) leží v
blízkosti lícního švu na vyvýšených
palpebrálních lalocích párové oči. Oči trilobitů mohou
být dvojího typu (obr. 3): holochroální
– početn očka leží vedle sebe a byla kryta
společnou rohovkou, schizochroální
– každé očko je kryto vlastní rohovkou.
Vývoj mnohých typů trilobitů vedl k redukci nebo
až k úplné ztrátě očí.
V takových případech došlo též
k vymizení lícních švů. Boční
okraje cephalonu často vybíhají do lícních trnů.
|
a |
b |
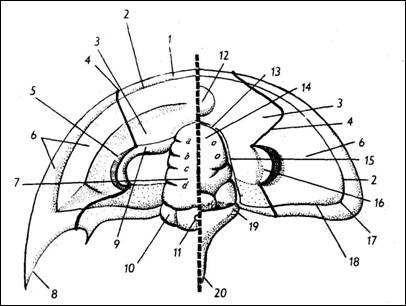

Obr. 2: a -Morfologie hlavového
štítu (podle Růžičky in Kumpera, Vašíček,
1986). 1 -
okrajová lišta, 2 - okrajová rýha, 3 - líce
pevné, 4 - lícní šev, 5 - zřecí plocha
oka, 6 - líce hybné, 7 - glabela, 8 - lícní trn, 9
- oční lišta, 10 - týlni lalok, 11 -
týlní hrbolek, 12 - čelní hrbolek, 13 -
předglabelární rýha, 14 - frontální
lalok glabely, 15 - rýha dorzální, 16 -
palpebrální lalok, 17 - lícní úhel, 18 -
zadní rýha, 19 - týlní prstenec, 20 -
týlní trn; u až d - glabelární rýhy. b
– hlavový štít rodu Paradoxides (foto P.
Skupien).
|
1 |
2 |


Obr. 3: Oči
trilobitů. 1 – holochloární oko rodu Asaphus, 2 –
schizochroální oko rodu Odontochile
(Clarkson, 1998).
Trup se skládá ze 2
až 44 pohyblivě skloubených článků.
Pohyblivé spojení článků umožňuje
stáčení (volvaci)
krunýře (obr. 4). Trupové články se
skládají z osy a
postranních pleur,
nesoucí obvykle hluboké pleurální rýhy. Pleury bývají na
povrchu hladké, případně zrnité.
|
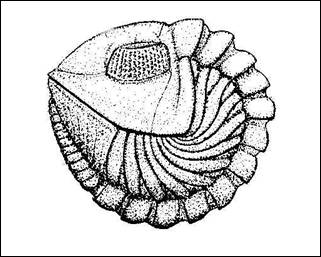
Obr. 4:
Stáčení krunýře (Clarkson, 1998). |
Obr.
5: Morfologie ocasního štítu trilobitů (podle
Hupého in Kumpera, Vašíček, 1986). 1 - osní část,
2 - pleurální část, 3 - artikulační
ploška, 4 - mezipleurální rýha, 5 -
pleurální brázda, 6 - kaudální trn, 7 ;
pleurální trny, 8 - druhotné okrajové trny, 9 - limbuá;
n až f - osní prstence. |
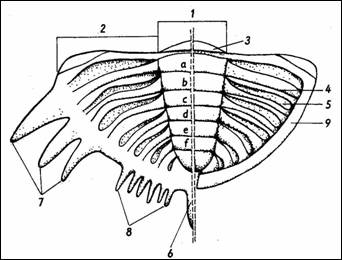
Pygidium (obr. 5) vzniklo splynutím
trupových článků a jsou na něm kromě
osní části (pygidiální osa) patrné pleurální rýhy i sutury na místě
splynutých článků (interpleurální rýhy). Utváření a
povrch krunýře je mimořádně různorodý,
s vráskami, hrbolky, trny, jamkami, terasovitými liniemi a
četnými dalšími strukturami.
Nálezy
celých skeletů jsou však vzácné. Je to
dáno malou odolností blanitých spojů mezi
částmi exoskeletonu. Po uhynutí jedince blanité spoje
podlehly rozkladu a proud nebo činnost jiných organismů vedla
k oddálení osamostatněných částí
krunýře. Při exuviaci došlo již primárně
k oddělení cephalonu a někdy volných lící
od zbývajících částí
krunýře. Běžně je pak nacházíme exuvie
splavené na jedno místo a velikostně
vytříděné.
Končetiny
trilobitů (obr. 6) byly článkované,
dvouvětevné, se silnější kráčivou
vnitřní větví (endopodit) a lupenitou
vnější větví (exopodit). Končetiny na cephalonu
nejsou zřetelně odlišeny od končetin trupu s
výjimkou prvního páru, který je
jednovětevný a přeměnen na tykadla (antény). Končetiny obdobně jako měkké
části se zachovávají zcela
mimořádně.
|
|
Obr. 6: Rekonstrukce končetin trilobitů: 1
– končetina pátého trupového
článku u rodu Triarthus, 2 – první
trupový článek u rodu Olenoides,
3 – druhý trupový článek u rodu Naraoia (Robison in Kvaček et
al., 2000). |
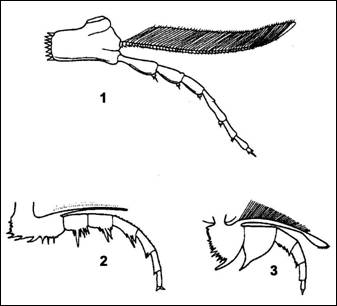
Trávicí
soustava začínala ústy ležícími na
spodní straně hlavy za hypostomem. Před ústy
neležely končetiny specializované k lovu nebo
přidržování potravy (např. jako u
korýšů). Jejich stavba je velmi podobná
končetinám na trupových článcích a
pygidiu. Předpokládá se, že jejich otrněné
kyčelní články mohly přidržovat a
případně i drtit ulovenou kořist. Jícen
pokračuje přímo do složitě větveného
žaludku v cephalonu. V ose těla vybíhalo ze žaludku
trubicovité střevo, které končilo řitním
otvorem na konci těla. Jiné orgány nejsou prokazatelně
doloženy.
Trilobiti byli
pravděpodobně odděleného
pohlaví, u některých druhů je popisován
pohlavní dimorfismus. Z vajíček se líhly larvy
(obr. 7). Počáteční larvální protaspidní stadium mělo krunýř rozdělený
pouze na cephalon a pygidium. Krunýř v tomto stadiu dosahuje
0,25 –
|
|
Obr. 7: Ontogenetický
vývoj rodu Sao. 1 –
raně protaspidní stadium, 2, 3 – pozdně
protaspidní stadium, 4 – raně meraspidní stadium, 5
– pozdně meraspidní stadium, 6 – dospělý
jedinec (Swinnerton in Clarkson, 1998). |
EKOLOGIE
Trilobiti žili výhradně v moři. Trilobiti byli převážně po dně nebo v blízkosti dna se zdržující bentózní živočichové, vyhledávající organický detrit a případně i lovící menší bezobratlé. Byli úzce přizpůsobení speciálním ekologickým nikám, v závislosti na charakteru sedimentu, hloubce, obsahu kyslíku ve vodě, klimatu a na dalších fyzikálně-chemických faktorech. Někteří, zejména velcí trilobiti byli i dravci. Stopy po predaci trilobitů jsou sice doloženy, jasný není způsob usmrcení a pozření větší kořisti, neboť trilobitům chyběly specializované končetiny k uchopení a rozdělení větší kořisti. Někteří trilobitů filtrovali organický detrit z usazenin na dně a vody nízce nade dnem, některé skupiny žily zahrabány do sypkého dna. Někteří trilobiti byli slepí, žili zahrabáni ve dně nebo uvnitř dutin, popřípadě dávali přednost hlubokovodním podmínkám, kde byl již nedostatek světla (obr. 8). Drobní a středně velcí trilobiti s nápadně tenkými krunýři jsou považováni za zástupce planktonu a nektonu. Trilobiti byli obecně horšími plavci, jsou však i výjimky. Trilobiti osídlovali mělkovodní litorální prostředí až prostředí hlubšího šelfu.
|
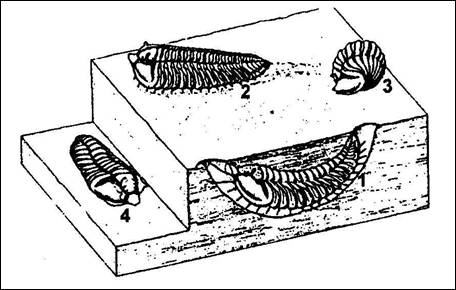
|
Obr. 8: Různé
životní pozice rodu Neseuretus.
1 – předpokládaná pozice v doupěti, 2
– lezení po dně, 3 – stočený jedinec, 4
– mrtvý jedinec (Hammann, |
Stratigrafie
Nejstarší
trilobiti se objevují ve spodní
kambrium. V průběhu kambria převládají typy s
vysokým počtem trupových článků. Na konci
kambria většina skupin vymírá a do ordoviku
přežívá jen několik skupin. Ve spodním
ordoviku však prodělávají rychlou adaptivní
radiaci a osídlují nová prostředí,
zejména prostředí útesová. Další
diverzifikace probíhá ve středním a svrchním
ordoviku, kdy trilobiti dosahují vrcholu rozvoje a
největší morfologické pestrosti. V průběhu
siluru a devonu zůstávají významnou skupinou bentosu.
Od konce devonu, po vymírání na rozhraní
středního a svrchního devonu, jsou již jen okrajovou
skupinou bezobratlých. V karbonu a permu jsou málo zastoupeni. Na
konci permu skupina vymírá úplně.
Trilobiti jsou
významnými vůdčími fosiliemi. Kromě
biostratigrafické zonace (např. v kambriu) se trilobiti
využívají i k vymezení faunistických paleoprovincií.
Např. v ordoviku lze rozlišit calymeno-dalmanitidní chladnovodní provicii
(Francie, severní Afrika, střední Evropa, Turecko), bathyuridní provincii v tropickém
klimatu (Severní Amerika, Sibiř a severní Čína),
asafidní provincii v
subtropickém klimatu Baltoskandinávie a Ruské platfomy a dikelokefalidní provincii v jižní
Číně a Autrálii, tehdy v mírném
pásu Gonwanského kontinentu.
SYSTÉM
Systematické
rozdělení trilobitů je založeno na morfologii
krunýře.
Řád Redlichiida
(spodní
- střední kambrium)
Výchozí
skupina trilobitů s velkým polokruhovitým cephalonem a
dlouhými lícními trny, četnými
trupovými články vybíhajícími do
trnů a drobným pygidiem. Oči jsou velké.
Eccaparadoxides, Paradoxides (obr. 9).
|
|
Obr. 9: Paradoxides
sp., kambrium, Podmokelský Mlýn. (foto P.
Skupien) |

Řád Agnostida
(spodní
kambrium - svrchní ordovik)
Drobní
trilobiti s podobně vypadajícím cephalonem a pygidiem. Jsou
slepí, s typickým diskovitým stočením.
Peronopsis
Řád Corynexochida
(spodní
kambrium - střední devon)
Heterogenní
řád s glabelou různého tvaru, obvykle s
rovnoběžnými okraji nebo
rozšířující se anteriorně.
Lícní šev opistopární. Pygidium je
velké, trup ze sedmi nebo osmi článků.
Ectillaenus,
Illaenus, Radioscutellum, Scutellum.
Řád Lichida
(střední
kambrium - střední devon)
Krunýř
je obvykle otrněný nebo s granulemi. Pygidium je
krátké, silně otrněné.
Odontopleura
Řád Phacopida
(spodní
ordovik - svrchní devon)
Výrazně
rozšiřující se glabela, schizochroálníma
očima, propárním lícním švem a nepřítomností
rostra. Trup z 11 článků. Pygidium s četnými
žebry, středně velké nebo dosahující
velikosti cephalonu.
Ananaspis, Dalmanitina (obr. 10) Odontochile,
Phacops.
|
|
Obr. 10: Dalmanitina sp., devon, Beroun. (foto P.
Skupien) |

Řád Ptychopariida
(spodní
kambrium - svrchní devon)
Trilobiti s
četnými trupovými články, menším
pygidiem a nečleněnou i členěnou glabelou Často
mají redukované oči nebo jsou slepí.
Conocoryphe, Ellipsocephalus (obr. 11), Ptychoparia, Sao, Harpes.
|
|
Obr. 11: Ellipsocephalus
hofii, kambrium, Jince. (foto P. Skupien) |

Řád Asaphida
(svrchní
kambrium - silur)
Menší
trilobiti s krátkým plochým trupem z menšího
počtu článků a cephalonem s velkou nečleněnou
glabelou. U většiny rodů je vyvinut perforovaný lem okolo
cephalonu, vytažený do dlouhých lícních
trnů.
Cryptolithus, Marrolithus.
ZPĚT NA systém
ZPĚT NA ÚVODNÍ STRANU