7.
Paleozoikum
Fanerozoikum (eonotém; období zjevného života) je tvořeno třemi eratémy: palezoikum, mezozoikum a kenozoikum. Paleozoický eratém má nejdelší trvání ve fanerozoiku, jeho rozpětí je přibližně 300 miliónů let. Paleozoikum se dělí na 6 útvarů, z nichž kambrium, ordovik, silur a devon jsou označovány jako starší (spodní) paleozoikum. Zbylé dva útvary karbon a perm jako mladší (svrchní) paleozoikum. Spodní paleozoikum započalo téměř na všech kontinentech postupnou mořskou transgresí (mořská záplava) v nízko ležících pobřežních oblastech. Konec paleozoika se vyznačuje celosvětovým ústupem moře.
Superkontinent Rodinia se začal rozpadat během proterozoika. Gondwana jako největší kontinent zahrnovala bloky jižní Ameriky, Afriky, Austrálie, Antarktidy, Přední Indie, Arábie i dnešní jižní Evropy. Mezi další důležité kontinenty patřil laurentský (převážná část Severní Ameriky), baltický (severní Evropa), sibiřský, kazašský a dva čínské kontinenty. Přetrvávající kadomská orogeneze počátkem paleozoika utichá. K významným procesům paleozoika patří kaledonská a variská orogeneze, která měla za následek opětovný vznik nového superkontinentu Pangea. Tento geotektonický cyklus započal rozpadem jednotného superkontinentu a byl ukončen jeho opětovným utvořením. Doprovodným procesem byla tvorba pásemných pohoří. Paleozoikum je známé bohatým rozvojem fauny i flóry. Hraniční interval proterozoika a paleozoika má stanovený stratotyp v jihozápadní části poloostrova New Foundland. Tato hranice je zde definována na bázi ichnofosilní stopy priapulidních červů Phycodes pedum (Trichophycus).
|
|
|
Obr. 7.1 Vymezení paleozoika. |
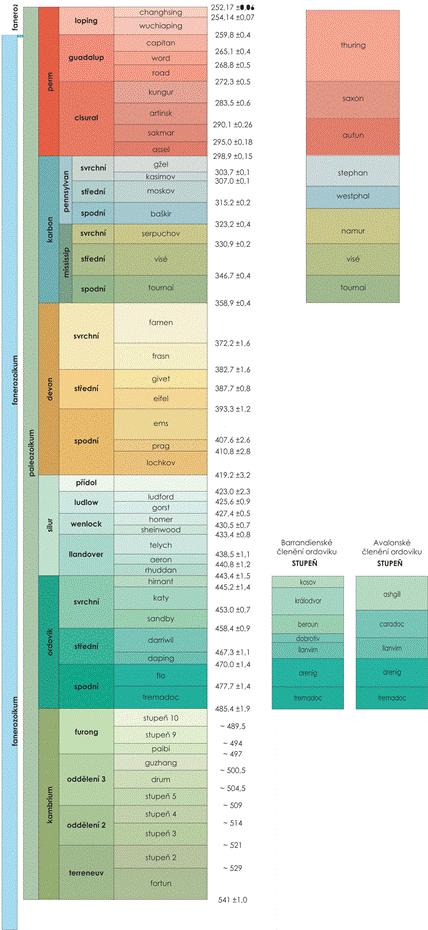
7.1
Kambrium (541,0 ± 1,0 až 485,4 ± 1,9 milionů let)
Nejstarší útvar paleozoika - kambrium trvalo zhruba 56 milionů let. Název kambrium pochází z latinského označení Walesu - Cambria a byl poprvé použit v 19. století Sedgwickem pro spodní část vrstevního sledu ležícího diskordantně nad prekambriem.
Na počátku kambria probíhal pokles pevninských bloků a následná transgrese, která dosáhla maxima uprostřed kambria. Transgrese nejvíce postihovala severní polokouli, na rozdíl od jižní polokoule, kde byla Gondwana skoro po celou dobu vynořená. Na počátku kambria doznívalo chladnější podnebí. Oteplení nastalo až ke konci kambria, kdy došlo k ustoupení moře a zvětšení souší.
|
|
|
Obr. 7.1.1 Relativní klimatické změny v historii Země. |
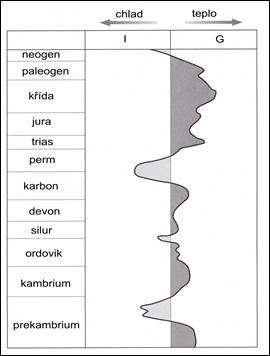
|
|
|
Obr. 7.1.2 Slepence spodního kambria, Čenkov. |

Pozůstatky flóry z kambria jsou zanedbatelné. Nejhojnější nálezy patří pozůstatkům různých řas. Suchozemské rostlinstvo není známo, jen malé části objevů se připouští suchozemský původ. Primitivní rostliny pozvolna pronikaly na souš. Vlivem neustálé transgrese a ustupování moří docházelo ke kolísání vodní hladiny a tehdejší ústrojenci (sinice a řasy) se dostali na souš do vzdušného prostředí. Probíhala invaze života na pevninu. Velká část se těmto změnám nedokázala přizpůsobit a hynula, ostatní se dokázali přizpůsobit např. tvorbou odolných výtrusů a nevlídné podmínky přežili. Vznikly tak první rostlinné obojživelné typy. Ve vodě přijímaly rostliny živiny celým povrchem těla a nepotřebovaly pevnou stavbu, na souši tomu však tak nebylo, museli mít pevnou pokožku, která je ochraňovala před odpařováním vody. Tělo rostlin bylo obohaceno o průduchy, které umožňovaly přijímání oxidu uhličitého a vydávání kyslíku. Dále bylo tělo vyztuženo tlustostěnným podpůrným pletivem, postupem času se vyvinulo i vodivé pletivo (cévice, později cévy). K přežití na souši byla také potřebná úrodná půda, čemuž napomohla činnost mikroorganismů i následný rozklad jejich odumřelých těl.
Kambrium bylo ve znamení bouřlivé exploze života (kambrická exploze života). Průvodcem této exploze byla rostoucí koncentrace kyslíku v ovzduší, dále také prokysličení hlubších partií vod oceánů a snížení intenzity ultrafialového záření. Zvýšení obsahu kyslíku se projevilo také ve zvýšené rychlosti životních pochodů. Fauna kambria zastupuje téměř všechny kmeny bezobratlých, včetně prvních zástupců obratlovců. Tomuto rozsáhlému stupni života v kambriu předcházel dlouhý vývoj v prekambriu.
Kdybychom měli možnost nahlédnout pod hladinu kambrického moře, byli bychom určitě velice překvapení, jaký pestrý život by nás tam čekal. V těchto mořích přibližně před 570 miliony lety probíhal velmi poklidný život. Většina obyvatel jak rostliny, tak i většina živočichů žila přímo na mořském dnu. Žádné ryby se v těchto mořích ještě nevyskytovaly.
Za nejvýznamnější skupinu kambria jsou považováni bez pochyb trilobiti, patřící do kmene členovců (Arthropoda). Během kambria trilobiti dosáhli mocného rozvoje, rozrůznili se do velkého množství skupin i druhů. Trilobiti díky svému nevšednímu tvaru a dobrému zachování ve fosilním stavu patří k symbolu paleontologie. Jsou velmi dobře známí i lajkům, a pro sběratele mají nesmírnou hodnotu. Jejich předkové se patrně vyvinuli z jednoduchých mořských kroužkovitých červů. Během 300 milionů let, kdy přežívali na této Zemi, vytvořili nespočetné množství rozmanitých druhů. Jednotlivé druhy trilobitů se lišily jak ve velikosti, tak ve tvaru těla či způsobu života. Trilobiti měli schránku složenou z celé řady článků, které byly vzájemně propojeny a přizpůsobeny k pohybu. Krunýř plnil funkci ochrannou, byl pevný a zároveň i ohebný. Během svého života trilobit krunýř několikrát svlékal z důvodu jeho růstu a stavěl si krunýř nový, větší. Tomuto procesu svlékání krunýře se říká exuviace, ta je významná pro znásobení nalezených materiálů, které podléhají fosilizaci.
|
|
|
Obr. 7.1.3 Kambričtí trilobiti (v popředí rod Paradoxides gracilis, v pozadí rod Conoryphe sulzer) a medúzy rodu Protolyella (autor obrázku Z. Burian). |

|
|
|
|
Obr. 7.1.4 Ellipsocephalus hoffi, kambrium, Jince. |
Obr. 7.1.5 Paradoxides gracilis, kambrium, Jince. |


Hojné také byly různé skupiny ramenonožců (Brachiopoda), které tvořily téměř třetinu obyvatel tehdejších moří, a stále jich přibývalo. Ramenonožci pomocí dlouhého stvolu byli přichyceni k mořskému dnu. Další hojnější skupinu kambria tvoří ostnokožci (Echinodermata). Stonek držící vakovitý kalich s rameny byl pevně zakotven kořenovým systémem na písčitém dně mělkých moří. Rozšíření byli také v kambrických mořích archeocyáti (Archaeocyatha) s typickou kornoutkovitou kostrou, kteří zastávali dřívější roli korálů. Jejich kalíškovité kostry vytvářely mohutné vápencové útesy dokazující, že moře v těchto oblastech bylo mělké a teplá.
|
|
|
Obr. 7.1.6 Vápenec tvořený zástupci rodu Archaeocyathus, spodní kambrium, Sibiř. |

Měkkýši (Molussca) a břichonožci (Gastropoda) jsou zastoupeni jen vzácně. První zástupci hlavonožců (Cephalopoda) byli prokázáni ze svrchního kambria ve východní Asii. Kambrická fauna se držela na okrajích mořských pánví a obývala jen mělká moře, kde se dokázala přizpůsobit nejrůznějším životním podmínkám (salinita, teplota vody, mořské proudy). Fauna byla tedy omezena výhradně na moře. Druhy, které by bylo schopny přijímat atmosferický kyslík, zatím nebyly vyvinuty. Tempo života se ovšem pomalu zrychlovalo a začaly vznikat předpoklady pro osídlení souše.
Stratigraficky velmi významnou spodnopaleozoickou skupinu představují Graptolithina náležející do kmene Hemichordata. V kambriu jsou zastoupena skupinou bentózních a pseudoplanktonních koloniových forem (Dendroidea).
Vedle skeletálních zbytků však v kambriu nacházíme i fauny s neobvykle četnými otisky živočichů s měkkým tělem. Jsou označovány jako fauny burgesského typu podle světoznámé středněkambrické lokality u Burgess Pass (Burgesského průsmyku) v Britské Kolumbii v Kanadě. V burgesské fauně se objevují i strunatci. Ještě během nejvyššího kambria se setkáváme s kožními kostmi či deskami bezčelistnatců (Agnatha) a v siluru s čelistmi (Gnathostomata).
|
|
|
Obr. 7.1.7. Bohatý život kambrického moře (autor Z. Burian). |

Z hlediska eustatických pohybů mořské hladiny je pro spodní
kambrium příznačný transgresní trend, který vrcholí ve středním kambriu.
Laurentie, sibiřský a kazašský kontinent leží na začátku kambria zhruba v
rovníkové oblasti a díky vysoké hladině světového oceánu jsou téměř úplně
zality teplým tropickým mořem. Konec kambria se vyznačuje regresí. Horotvorné
pochody byly ve znamení doznívání
kadomské orogeneze. Intruze plutonitů byly též doprovázeny suchozemským
výlevným vulkanismem.
|
|
|
Obr. 7.1.8 Ryolity v lomu Těškov, svrchní kambrium. |

|
|
|
Obr. 7.1.9 Poloha kontinentů v období raného kambria (upraveno podle Faupla, 2000). Zkratky: Ba – Baltika, Sv – Špicberky, Ka – Kazachstán, Sč – Severní Čína, Jč – Jižní Čína, Ti – Tibet, Ar – Arábie, Alp – alpinská Evropa, Va – evropské variscidy, Sp – Iberský poloostrov. |
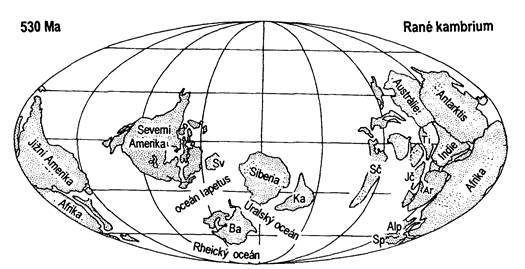
Během kambria se od sebe vzdalují Laurentie a Baltika, a prostor mezi nimi je vyplněn protoatlantickým oceánem neboli oceánem Japetus. Mezi Baltikou a Gondwanou existuje další oceánský prostor, oceán Paleotethys. Mezi Baltikou a Sibiří je otevřen uralský oceán. Vzdalování Laurentie, Baltiky a Gondwany přetrvává až do ordoviku a poté se na okrajích Japetu a Paleotethys začínají tvořit subdukční zóny a oceány se začínají uzavírat.
7.2
Ordovik (485,4 ±1,7 až 443,8 ± 1,5 milionů let)
Název ordovik poprvé použil Angličan Lapworth v 19. století. Název je odvozen od latinského označení keltského kmene Ordoviciů, kteří žili v severním Walesu.
Minimálně stejně bohatý, možná i o něco pestřejší život jak v kambriu, přinesl následující ordovik. Spodní hranice ordoviku je vymezena bází zóny dendroidních graptolitů Dictyonema flabelliforme, svrchní hranice je definována bází graptolitové zóny Akidograptus acuminatus. Ordovik se člení na tři epochy- spodní, střední a svrchní ordovik. V mezinárodním členění spodní ordovik tvoří stupně – tremadok a flo, střední ordovik – daping a darrivill a svrchní ordovik – sandby, katy a himant.
V běžné praxi se dosud používají britské stratigrafické stupně, které nejsou podloženy stratotypy a ve vyšším ordoviku nejsou podloženy ani dostatečně hojnou, vhodnou faunou. Havlíček a Marek (1973) se proto na základě uloženin v Barrandienu pokusili pro vyšší ordovik definovat české stratigrafické stupně. Ty jsou však dnes považovány jen za regionálně platné. Vedle graptolitů se v detailní biostratigrafii uplatňují rovněž konodonti.
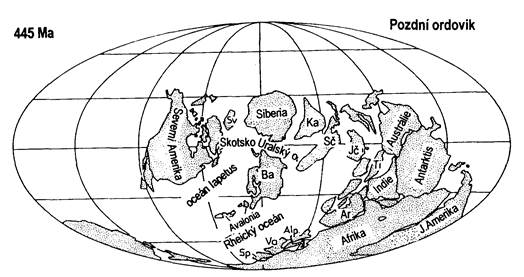
Začátkem ordoviku bylo rozložení kontinentů podobné jako v kambriu. Superkontinent Gondwana se stáčel a jižní část se nacházela až v oblasti jižního pólu. Ordovické klima bylo nestálé a na rozdíl od kambria byla běžná chladná období, která měla vliv i na stav světové hladiny oceánů. Pro počátek ordoviku jsou typické transgrese, které se střídaly s epizodickými regresemi (ústupy moře). Hladina oceánu se postupně zvedá až do svrchního ordoviku, kdy kulminuje v úrovni o několik set m vyšší než je hladina dnešní (až o 600 m?!). Nejvyšší ordovik charakterizují rozsáhlé globální regrese, doprovázené prudkým ochlazením. Zaledněná byla jižněji položená část Gondwany. Klimatické změny ovlivňovaly i rozvoj života.
|
|
|
Obr. 7.2.1 Poloha kontinentů koncem ordoviku. |
Nálezy flóry z ordoviku jsou dosti vzácné. Z našeho území byly popsány první cévnaté rostliny Boiophyton pragense a Kreiciella, které se považují za první rostliny suchozemské. Početnější zbytky vyšších rostlin bohužel nemáme, ale i přesto lze pokládat ordovik za období, kdy se jejich vznik utvářel. Tento vývoj byl zřejmý v siluru, kde již nalézáme dobře zachované pozůstatky skupiny Psilophyta. Přechodné typy mezi řasami a cévnatými rostlinami se ovšem nedochovaly.
Stejně jako v kambrických, tak i v ordovických mořích byli dominantou trilobiti, kteří zde dosáhli maximálního rozvoje. Těla trilobitů prošla určitými změnami, došlo ke zkracování těla, zvětšování ocasního štítu, na povrchu se začaly objevovat různé výrůstky ve formě trnů. Objevují se trilobiti velkých forem, kteří dosahovali až několik desítek centimetrů.
Ordovik charakterizují hojné výskyty ramenonožců. Jejich bílé vápnité schránky připomínající mlže, byly četně rozprostřeny po mořském dnu. Celosvětového rozšíření dosáhli graptoliti, kteří představují vymřelou třídu polostrunatců (Hemichordata). Graptoliti žijící v koloniích vylučovali organickou podpůrnou a ochrannou vnější kostru. Graptoliti se nejvíce zachovali v tmavých břidlicích, kde tvoří pilovitě vrubované nárysy. Žili v mořích na celém světě, protože se dokázali rychle přizpůsobit danému prostředí. Vytvářeli velké množství druhů, tyto druhy lze nalézt ve vrstvách stejného stáří na místech i tisíce kilometrů od sebe vzdálených. Na základě těchto druhů je možné dobře charakterizovat vrstvy stejného stáří, a je na nich založena stratigrafie a rozdělení ordoviku a siluru na celém světě.
|
|
|
Obr. 7.2.2 Obolus sp., ordovik, Barrandien. |

|
|
|
|
Obr. 7.2.3 Černé peitické sedimenty s graptolity. |
Obr. 7.2.4 Sedimentární hematit, spodní ordovik, arenig, Ejpovice. |


|
|
|
Obr. 7.2.5 Graptolitová břidlice. |

Měkkýši byli taktéž hojně zastoupeni. V ordoviku nastal jejich první větší rozvoj. Mlže již zastupovaly všechny základní řády. Běžně se vyskytují loděnkovití hlavonožci s dlouze protaženými kornoutovitými schránkami o velikosti až metrových délek. Loděnky patřili mezi dobré plavce. Kolem ústního otvoru měli několik desítek chapadel, před kterými bylo okolí v pozoru. Patřili totiž mezi první dravce. Hojní byli také v ordovických mořích ostnokožci (Echinodermata), nejčetněji zastoupeni skupinou jablovců a lilijic.
|
|
|
|
Obr. 7.2.6 Aristocystites bohemicus, ordovik. |
Obr. 7.2.7 Echinosphearites sp., ordovik. |


V ordoviku již nacházíme důkazy života významného podkmenu obratlovců (Vertebrata). Nálezy tvoří drobné kosterní destičky pocházející z rozpadlých krunýřů rybovitých praobratlovců, kteří ještě neměli vyvinuté čelisti (Agnatha). Tělo bezčelistních obratlovců bylo dlouhé zhruba 10–40 cm a bylo chráněno pevnými kostěnými deskami. Nálezy pozůstatků těchto obratlovců vyvolalo otázku, z které skupiny organismů se vyvinuli. Poslední studie poukazují na to, že se obratlovci pravděpodobně vyvinuli z polostrunatců (mladší vývojová skupina červů) a ostnokožců.
|
|
|
Obr. 7.2.8 Ordovické moře (autor obrázku Z. Burian). |

Rozložení kontinentů počátkem ordoviku odpovídalo situaci v kambriu. Superkontinent Gondwana se během ordoviku postupně stáčel tak, že jeho jižní části zasahovaly do jižní polární oblasti.
Počátkem ordoviku převládal transgresní režim. Sedimentace nejvyššího ordoviku probíhala ve znamení globální regrese, spojené s prudkým ochlazením a výrazným poklesem oceánské hladiny. Mořská sedimentace v ordoviku byla často doprovázená vulkanickou činností, jejímž produktem byly hlavně vyvřeliny bazaltového typu.
V období od ordoviku do spodního devonu dochází k významným přesunům kontinentů, které jsou provázeny horotvornými fázemi tzv. kaledonské orogeneze. Kontinenty Laurentie a Baltika se přibližují a mezi nimi se postupně uzavírá oceán Japetus.
7.3
Silur (443,7 ±1,5 až 416,0 ± 2,8 milionů let)
Pojem silur použil poprvé anglický paleontolog Murchison v roce 1835 při studiu geologických poměrů ve Walesu. Odvodil ho od jména keltského kmene Silurů.
Silur je s devonem jediným obdobím geologické minulosti, které je rozděleno na chronostratigrafické jednotky, které jsou všechny podložené stratotypy. Silur se dělí na oddělení llandover (se stupni rhuddan, aeron a telych), wenlock (se stupni sheinwood a homer), ludlow (se stupni gorst a ludford) a přídolí (bez dalšího dělení). Spodní hranice siluru je stanovena bází graptolitové zóny Akidograptus acuminatus, svrchní hranice na bázi graptolitové zóny Monograptus uniformis. Hranice jednotek jsou vymezeny podle změn v graptolitových faunách.
|
|
|
Obr. 7.3.1 Mezinárodní stratotyp hranice silur/devon, Klonk u Suchomast. |

Kontinenty v období siluru se nacházely poměrně blízko sebe a byly obstoupeny mohutným oceánem Panthalassa. Na jižní polokouli v blízkosti pólu se nacházel kontinent Gondwana. V blízkosti pólu na Gondwaně přetrvávalo chladné klima, dokonce i zalednění. Skupina menších kontinentů obklopovala rovník. Na západě byla Laurentia, na jihu malá Avalonie a na východě ležela Baltika. Všechny tyto kontinenty se pomalu přibližovaly k sobě. Koncem siluru došlo ke srážce Laurentie a Avalonie s Baltikou (zanikl oceán Japetus), proběhlo kaledonské vrásnění (mladokaledonská orogeneze), které později vedlo ke vzniku kontinentu Laurussie (případně též jako Old-Red kontinent - kontinent starého červeného pískovce podle charakteristicky zbarvených sedimentů). Vzniklo rozsáhlé kaledonské pásemné pohoří s bivergentní příkrovovou stavbou. Orogeneze je spojená s metamorfními účinky a s magmatickou činností. Některé oblasti, včetně Českého masívu, však ležely mimo dosah uvedeného vrásnění.
|
|
|
Obr. 7.3.2 Poloha kontinentů ve středním siluru. |
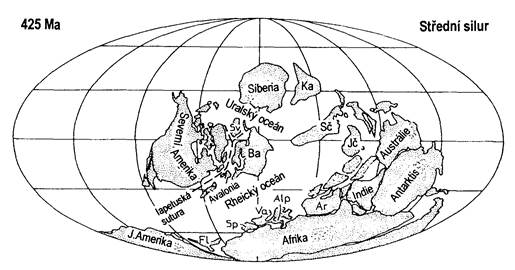
Po svrchnoordovickém silném ochlazení, které vedlo k vymírání, nastalo teplé silurské období doprovázené projevy skleníkového efektu. Na začátku siluru rychle ustupovaly ledovce a během středního siluru se téměř žádné nezachovaly. Světový oceán vlivem transgrese (tzv. hraniční ordovicko-silurský event.) ve spodním siluru zaplavil ploché pobřežní kontinenty, čímž vznikla rozsáhlá šelfová moře, charakteristická celosvětovou sedimentací anoxických graptolitových břidlic. Předpokladem této plošně rozsáhlé anoxie v poměrně mělkých podmínkách (v oblastech hlubšího šelfu) je, že došlo k výstupu málo okysličených vod z hloubek během obnovení cirkulace vodních mas při vyrovnání teplotních rozdílů svrchních (zamrzlých) a spodních (chladných) vrstev oceánu, popřípadě došlo k míšení šelfových moří se sladkou vodou pocházející z tajících ledovců. Výsledkem těchto procesů byla stabilizace celosvětového klimatu, na rozdíl od nestálého ordoviku. Časem anoxie vyzněla, a začaly se objevovat stále vyspělejší druhy mělkovodní fauny.
|
|
|
|
Obr. 7.3.3 Graptolitové břidlice, lom Kosov. |
|


Po celkovém oteplení, kdy byly mělké vody dobře prohřáté, a prokysličené se začalo dařit korálnatcům (Anthozoa). Korálnatce představuje skupina koloniálních deskatých korálů (Tabulata) a skupina korálů drsnatých (Rugosa), kteří žili samostatně. Kromě korálnatců byla v období siluru běžná skupina stromatopor (Stromatoporoidea), která vylučovala pevné vápnité kostry bochníkovitých útvarů dosahujících až 2 m v průměru. Korálnatci a stromatopory se staly základem četných silurských vápenců. V těchto silurských vápencích se často objevují schránky měkkýšů (Mollusca), u nich lze v siluru pozorovat značný rozvoj. Měkkýši představovali hlavní složku silurského moře. Mezi nejběžnější měkkýše v siluru patří loděnky (Nautiloidea), ty mohly dosahovat ohromných rozměrů až 4,5 m. Loděnky se postupem času vyvíjely, nejprve měly schránku rovnou, později se začaly schránky stáčet. Zajímavostí je loděnka hlubinná (Nautilus pompilius), která jako žijící fosilie nevymřela a dodnes se objevuje v tichém oceánu. V silurském moři se život stále ještě držel především u dna. Husté porosty tvořené lilijicemi a korály, poskytovaly úkryt drobným tvorům. Lilijice (Crinoidea, kmen Echinodermata) byly přichyceny dlouhým stonkem k mořskému dnu a na opačném konci se nacházelo tělo živočicha podobné lilii. V mělkých zálivech silurského moře bychom mohli spatřit pestrý život plný mnoha druhů zelených a červených řas, které pocházejí již z ordoviku. Místa, kde docházelo k mísení sladké a slané vody byla ideální pro hojné rybovité praobratlovce (Agnatha), kteří se pomalu pohybovali nad dnem. Graptoliti jako běžní obyvatelé silurských moří měli stejně jako v ordoviku z hlediska stratigrafie obrovský význam, slouží k přesnému srovnávání silurských vrstev na celém světě. Láčkovci (Coelenterata) dosáhli taktéž značného rozvoje. Bohužel ne u všech živočichů tak jak u výše zmíněných probíhal rychlý rozvoj, některé skupiny v rozvoji zaostávali, dokonce i pomalu vymírali. Do této skupiny patří například některé čeledi trilobitů a ostnokožci.
|
|
|
Obr. 7.3.4 Halysites sp., Kozolupy. |

|
|
|
|
Obr. 7.3.5 Ramena lilijice Scyphocrinites elegans, silur, články stonku lilijic rodu Scyphocrinites, silur, Barandien |
|


Již z ordoviku známe primitivní skupinu bezčelistných (Agnatha), ke které se v siluru připadali zástupci obratlovců čelistnatých (Gnathostomata) s vyvinutou spodní i svrchní čelistí. Z této skupiny se objevují ryby trnoploutvé (Acanthodii), které měly hlavní rozvoj především v pozdějším devonu.
Souše zatím byly pusté, bez známky života. Pouze ve vlhčích místech, například v bažinách, v okolí jezer, nebo v deltách řek bylo možné pozorovat občasnou vegetaci. Na souších se objevují nejstarší cévnaté (suchozemské) rostliny.
|
|
|
|
Obr. 7.3.6 Cooksonia sp. |
|


|
|
|
Obr. 7.3.7 Silurské moře v době před 400 miliony let v oblasti dnešních středních Čech. |

7.4 Devon (416,0 +- 2,8 až 359,2 +- 2,5 miliónů let)
Devon definovali dva význační angličtí geologové Sedgwick a Murchison. V roce 1839 zjistili, že fosilie tohoto útvaru mají přechodný charakter mezi silurem a karbonem a nový útvar pojmenovali podle hrabství Devonshire na jihozápadním pobřeží Anglie.
Stratotypem spodní hranice
devonu (současně celého devonu a prvním stanoveným stratotypem na světě) je
profil na Klonku u Suchomast poblíž
Berouna. Devon se dělí na spodní (stupeň
lochkov, prag, ems) střední (stupeň eifel, givet) a svrchní
(stupeň frasn, famen). Spodní
hranice je definována na základě prvního výskytu graptolita Monograptus uniformis. Svrchní hranice
s karbonem má stratotyp v jižní Francii v La Serre v pohoří
Montagne Noire a je stanovena prvním výskytem konodonta Siphonodella sulcata.
Nejstarší stupně devonu (lochkov a prag) jsou definovány stratotypy v Barrandienu, následující stupeň ems ve střední Asii. Ems je v Barrandienu rozdělen na nižší část - zlíchov a na vyšší část - dalej. Jejich mezinárodní uznání na úrovni stratigrafických stupňů je zatím předmětem jednání. Eifel je definován v Rýnském břidličném pohoří, givet v Maroku. Svrchnodevonské stupně frasn a famen a hranice s karbonem mají své stratotypy ve Francii v pohoří Montagne Noire.
Devon je považován za neklidné období. V první polovině zcela utichlo kaledonské vrásnění. Doznívají projevy mladokaledonského vrásnění, spojeného s pozdně orogenní plutonickou činností a s kyselým výlevným vulkanismem. Zhruba před 420 milióny lety byl zformován prvohorní kontinent Laurussie (vzniklý srážkou Laurentie a Avalonie s Baltikou). Laurussie je jinak známá jako Severoatlanský nebo Old Red kontinent. Tato devonská prapevnina „Starý červený kontinent“ angličtí geologové pojmenovali podle charakteristických sedimentů – červených pískovců. Byli zde objeveni první suchozemští čtvernožci, také primitivní obojživelníci (Amphibia). Ve středních Čechách v této době probíhala nepřetržitá mořská transgrese a došlo k nepřetržitému vývoji různých ústrojenců, proto je tato oblast považovaná za světový standard pro vymezení hranice mezi silurem a devonem. Většina kontinentů byla soustředěna v teplém klimatickém pásmu. V chladném pásmu se nacházely jižní části Gondwany, kde devon má siliciklastický vývoj (bez vápenců).
|
|
|
Obr. 7.4.1 Poloha kontinentů ve středním devonu. |
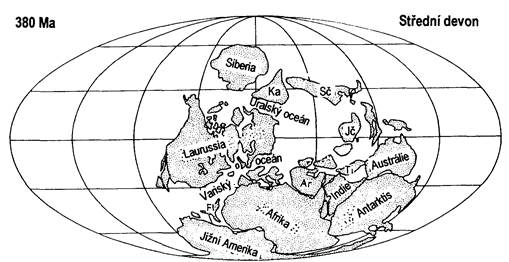
V nově formovaných sedimentačních prostorech od středního devonu nastupují procesy variského (hercynského) vrásnění. V některých se uplatňuje silný submarinní vulkanismus.
Podle nalezené flóry na starém červeném kontinentu přetrvávalo teplé a suché podnebí. Ke konci devonu přibývalo srážek. Následkem zvýšeného množství srážek se urychlil rozvoj cévnatých rostlin a vznikly první uhelné sloje. Na rozvoj života v devonu měl vliv nejen vznik starého červeného kontinentu, ale zejména vznik mělkých teplých moří s četnými zálivy ideálními pro rozvoj obratlovců, především ryb. Devon je označován jako období ryb. Na severní polokouli byly četné vodní plochy jako řeky, laguny, jezera a močály, a v těchto místech se dařilo rozvoji suchozemských rostlin. Zde se také vyvinuli první obojživelníci schopní dýchat vzduch.
Období devonu bylo velice příznivé pro rostlinstvo. V prvních suchozemských pralesech bylo možné vidět kapraďorosty i první typy stromovitých rostlin. Od samého počátku se jedná o první okamžik, kdy rostlinný vývoj předbíhá vývoj živočišný. Vznikaly stále nové úrodné oblasti, tím pádem stále nové rostlinné druhy. Z prvotních mořských řas, později vznikly bažinné a následně suchozemské rostliny bez přímého kontaktu s vodou. Devon přinesl ideální podmínky pro „dobytí“ pevniny rostlinstvem. Tyto pokusy probíhaly marně v přecházejících obdobích od kambria do siluru.
Nejstarší a nejdůležitější skupinou výtrusných devonských rostlin byla Psilophyta. Tato rostlina vzniklá z některého druhu řas (předpokladem je řasa zelená) byla maximálně 100 cm vysoká, nepravidelně větvená a na povrchu buďto holá nebo pokrytá trnovitými lístky. Mnoho rostlin a živočichů se neumělo přizpůsobit devonským horotvorným a klimatickým změnám a vymíralo. Největší klimatická změna svrchního devonu je označovaná jako kellwasserský event (na hranici stupně frasn a famen), kdy došlo k prudkému ochlazení, které mělo za následek vymření až 70% všech rodů fauny. Ostatní schopnější rostliny naopak vytvořily nové formy. V souvislosti s ním končí růst korálovo-stromatoporoidových útesů a dochází k výrazné redukci nebo k vymírání některých faunistických skupin. Někteří tuto výraznou událost vysvětlují impaktem nebeského tělesa.
|
|
|
Obr. 7.4.2 Fauna je tvořena různými typy přesličkovitými, plavuňovitými a kapraďovitými rostliny. |

Mělká, teplá moře byla v průběhu devonu stále plná drsnatých a deskatých korálů a také stromatoporů. Na konci devonu ovšem přichází zlom a rozvoj těchto hojných skupin pomalu ustupuje. Rozvoje dosáhli v devonu ramenonožci a některé skupiny měkkýšů a první drobní zástupci stočených hlavonožců ze skupiny amonitů (Ammonoidea). Amoniti jsou stratigraficky velice významní, a je možné díky nim porovnávat stejně staré vrstvy devonských sedimentů na celém světě. Břichonožci (Gastropoda) a mlži (Bivalvia) se vyskytovali pouze v primitivních formách. Mlži dokonce pronikali do sladkých vod.
|
|
|
|
Obr. 7.4.3 Actinostroma sp., devon, Koňeprusy. |
Obr. 7.4.4 Amphipora sp., devon, Macocha. |


|
|
|
Obr. 7.4.5 Sieberella sieberi, devon, Koněprusy. |

|
|
|
|
Obr. 7.4.6 Fenestella sp., devon, Beroun. |
Obr. 7.4.7 Utropora nobilis, devon, Beroun. |


Jak už bylo výše zmíněno, devon patřil rybám. Vody byly přeplněné rybami, které obývaly jak slanou, tak i sladkou vodu. Z devonských rybovitých obratlovců známe třídy praobratlovců (Agnatha), čelistnatých ryb (Gnathostomata), pancířnatých ryb (Placodermi), kostnatých ryb (Osteichthyes), lalokoploutvých ryb (Crossopterygii), trnoploutvých ryb (Acanthodii), paryb (Chondrichthyes), paprskoploutvých ryb (Actinopterygii), dvojdyšných ryb (Dipnoi), řídkokostných ryb (Chondrostei), mnohokostných ryb (Holostei), celokostných ryb (Teleostei). Každá jednotlivá třída těchto rybovitých obratlovců měla pod sebou několik druhů, které dominovaly ve spodním devonu, ale bohužel většina z nich ke konci devonu vymírá.
V devonu proběhla důležitá událost, která měla na souši prvořadý význam a to ten, že z ryb vznikl první obojživelník. První obojživelníci byli téměř totožní s rybami lalokoploutvými. Otázkou je, proč vodu opouštěli a vylézali na souš. Poměrně paradoxním závěrem je, že přecházeli na souš, aby mohli zůstat ve vodě. Docházelo k vysychání močálu i jezer, a tak byli obojživelníci donuceni po souši se přesunout tam, kde voda ještě byla. K přelézání z jednoho močálu do druhého měly tyto přechodné formy vyvinuty primitivní plíce, díky kterým mohly dýchat kyslík. Dalším důvodem přechodu na souš bylo pravděpodobně to, že se rádi vyhřívali na slunci. Prvně navštěvovali prohřáté mělké pobřežní vody, občas vylezli na předměty trčící z vody, a když jim to bylo málo, posléze vylezli až na břeh, kde navíc měli na blízku i potravu. Jelikož tito první obratlovci neměli žádnou konkurenci, mohli se rychle rozmnožovat a tak vznikaly nové formy, které rychle pokryly souš.
|
Obr. 7.4.8 Na obrázku dravý Dunkleosteus, který dorůstal délky více jak 10 metrů a váhy až čtyř tun. |
|
Obr. 7.4.9 První přesuny po souši. |
Devon byl z hlediska rozvoje velice důležitým obdobím. Souš byla pokryta prvními pralesy a obojživelníky. Tento vývoj je následkem mohutné revoluce kaledonské, která zapříčinila změny na zemském povrchu. Život se musel těmto podmínkám přizpůsobit, a pokud to nezvládl tak zanikl.
|
|
|
Obr. 7.4.10 A:
pancířnaté ryby (Placodermi), druh Bothriolepis canadensis, B:
lalokoploutvé ryby (Crossopterygii),
druh Eusthenopteron foordi,
Zástupci hlavních skupin kostnatých ryb; C: Moythomasia nitida (paprskoploutvé ryby Actinopterygii), D: Scaumenacia curta (dvojdyšné ryby Dipnoi), E: Eusthenopteron foordi (lalokoploutvé ryby Crossopterygii), F: paprskoploutvé ryby (Actinopterygii), druh
Cheirolepis canadensis, G: mnohokostnaté ryby (Holostei), druh Dapedium
pholidotum, H: celokosté ryby (Teleostei),
druh Leuciscus, Zástupci
lalokoploutvých ryb (Crossopterygii),
ze skupiny střapcoploutvých I: Latimeria,
B: Macropoma, C: Coelacanthu. |

V evropském prostoru v teplém klimatickém pásmu se rozlišují tři základní vývoje devonu:
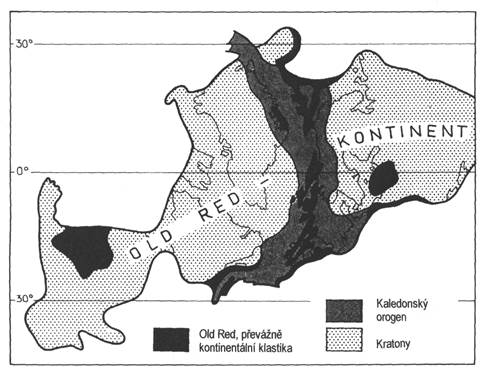
1) Vývoj starého červeného pískovce (Old Red). Charakterizují ho převážně klastické až hrubě klastické, pestře zbarvené uloženiny příbřežních lagun, jezer a řek vzniklé v kontinentálním prostředí (obr. 8). Hojné bývají suchozemské rostliny.
2) Vývoj český s převahou širokomořských (pelagických) uloženin. Hojné jsou karbonáty, ve fauně pelagické planktonní a nektonní organismy.
3) Vývoj rýnský s převahou mělkovodních sedimentů tvořených terigenním klastickým materiálem. Typické jsou plážové a deltové uloženiny. Ve fauně převládá bentos.
|
|
|
Obr. 7.4.11 Kontinent starého červeného pískovce ve spodním devonu. |
|
|
|
|
Obr. 7.4.12 Vápence devonu, Velká Amerika. |
Obr. 7.4.13 Moravský kras, Rudice. |


7.5
Karbon (359,2 +- 2,5 až 299,0 +- 0,8 miliónů let)
Svrchní paleozoikum tvoří dva útvary, karbon a perm.
Název karbon (“Carboniferous“ – angl. „uhelný“) byl zaveden anglickými geology Connybearem a Phillipsem již v r. 1822.
Spodní hranice karbonu je stanovena na základě prvního výskytu
konodonta Siphonodella sulcata a o
něco výše se nachází amonitová zóna Gattendorfia.
Spodní hranice je definována na stratotypu
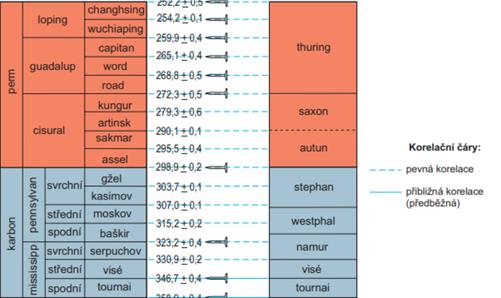
V západní a střední Evropě se uznává spodní a svrchní karbon jako oddělení. Dělení spodního karbonu v Evropě na stratigrafické stupně tournai a visé se běžně užívá a má naději na mezinárodní přijetí. Podstatně složitější je situace v karbonu vyšším, kde existují různé a vzájemně těžko kombinovatelné regionální stratigrafické stupně. Stratigrafické stupně svrchního karbonu, zavedené a užívané v západní a střední Evropě, namur až stephan slouží jen jako regionální chronostratigrafické jednotky.
|
|
|
Obr. 7.5.1 Chronostratigrafická tabulka permokarbonu. Levá strana – mezinárodní rozdělení, pravá strana - středoevropské rozdělení. |
Období karbonu charakterizují významné paleogeografické změny. Superkontinent Pangea se začal utvářet již během karbonu, a jeho vývoj byl dokončen v permu. Počátky karbonu vyznačují mocné horotvorné procesy označovány jako hercynské (variské) vrásnění. Kolize Gondwany s Laurusií vede k vyvrásnění variského horstva v Evropě a horského pásma při jihovýchodním a jižním pobřeží Severní Ameriky.
Dominantní pevnina Gondwana pokrývající velkou část jižní polokoule, byla obklopená mohutným oceánem Panthalassa. Gondwana v období karbonu byla z velké části zaledněná. Největší pevninou na severní polokouli byla Laurussie. Tyto dva kontinenty se k sobě stále přibližovaly. Kolize Gondwany s Laurussie vede k vyvrásnění variského horstva v Evropě a horského pásma při jihovýchodním a jižním pobřeží Severní Ameriky. V místě srážky vznikl mohutný systém horských pásem v délce 5000 km táhnoucí se podél rovníku. Pozůstatky v minulosti až několik kilometrů vysokých pásemných horstev lze vidět ve východní části Severní Ameriky (alleghenidy – pohoří Appalače), v severní Africe (mauretanidy – pohoří Atlas) a v Evropě (variscidy). V kolizní zóně se nacházel např. i mikrokontinent Armorika, k níž patřil Český masiv. Vrásnění v Evropě se dělí do jednotlivých etap: bretonská (na hranici devon-karbon), sudetská (na hranici spodní-svrchní karbon), krušnohorská (ve vyšším namuru a spodním westphalu) a asturská fáze (na hranici westphalu a stephanu).
|
|
|
Obr. 7.5.2 Poloha kontinentů ve spodním karbonu. |
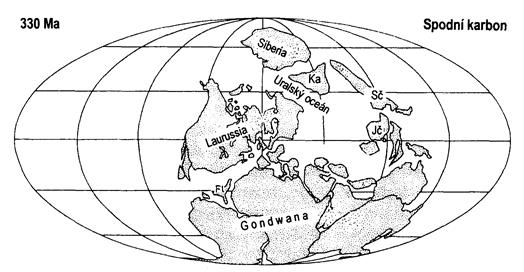
Důsledkem těchto horotvorných procesů bylo přerušení mořské sedimentace v oblasti středních Čech. Dalším doprovodným jevem byla značná sopečná činnost. Vznikaly mohutné žulové masivy (např. středočeský, krkonošsko-jizerský). Na okrajích četných lagun a při deltách řek se hromadilo velké množství rostlinné hmoty, které časem podléhalo chemickým procesům tzv. uhelnatění, a vznikala ložiska černého uhlí. Mořská hladina neustále kolísala a tak uhelné močály byly často zaplavovány. V období ústupu moře začaly vznikat nové pralesy. Tento proces se stále dokola opakoval. Ve variském pohoří se vytvořily vnitrozemské pánve s rozlehlými močály, kde nedocházelo k záplavám a ukládalo se tam kvalitní černé uhlí (okolí Plzně).
Klima v karbonu bylo proměnné. V počátcích v mississippu bylo podnebí teplé s vysokým obsahem CO2. Tento plyn pravděpodobně do ovzduší rozptýlila sopečná činnost. Značnou část spotřebovalo rostlinstvo a podnebí se měnilo stále v sušší. Ve svrchním karbonu bylo podnebí dokonce pouštní. Změny v podnebí jsou viditelné v usazených horninách, kdy během vlhkého podnebí se ukládaly vrstvy šedých hornin i uhelné sloje a během suchého podnebí se ukládaly vrstvy červených pískovců.
Hercynská revoluce je spojena s novým velkým rozvojem. Podnebí bylo vlhké, teplé s dostačujícím obsahem CO2, které je ideální pro rozvoj rostlinných druhů, často stromovitého růstu. Devonské stromovité plavuně reprezentují především dva nejvýznamnější druhy, a to Lepidodendron a Sigillaria. Rod Lepidodendron představoval plavuni až 30 m vysokou s bohatě rozvětvenou korunou obohacenou výtrusnými šišticemi. Pro Lepidodendron je typický povrch kmene, který byl pokryt listovými polštářky kosočtverečného tvaru s listovými jizvami.
|
|
|
Obr. 7.5.3 Kůra rodu Lepidodendron, Žacléř. |

|
|
|
Obr. 7.5.4 Větve plavuní, lokalita Žacléř. |

Druhým hojným
rodem stromovitých plavuní je Sigillaria.
Rod Sigillaria s výškou 20 m
patřil mezi hlavní rostliny evropských a severoamerických uhlotvorných
karbonských pralesů. Z jednoduchého kmenu vyrůstaly přesleny
s výtrusnicovými šišticemi. Naspodu se větvily kořeny známé pod označením
stigmarie. Kmen i větve se vyznačují jizvami po odpadlých listech.
Přesličky osídlovaly vlhké
prostředí. Většinou vyrůstaly z vod jezer či bažin. Přesličky dosahovaly
výšky okolo 20 m a jejich kmen s mohutnou dřeňovou dutinou mohl mít průměr
až 1 m. Listy přesliček byly úzké, čárkovité a rostly na pravidelně větvených
větvích. Stromovité přesličky reprezentuje především rod Calamites.
|
|
|
Obr. 7.5.5 Nákres přesličky. Obr 2. Calamites undulatus, svrchní karbon, OKR (sbírka VŠB-TU Ostrava). |


K dalším významným karbonským rostlinám patří kapradiny. Stromovité kapradiny obývaly spíše sušší místa a vyrůstaly do výše 10-15 m. Kapradiny vytvářely z listů objemné vějíře, které vyrůstaly buď jen ze dvou stran kmene, nebo z jeho celého obvodu. Část kapradin náležela pravděpodobně kapraďosemenným rostlinám (Pteridospermophytina), které se tvarem téměř nelišily od kapradin, avšak listy kapraďosemenných rostlin vytvářely skutečná semena. V karbonských pralesech se objevují i první primitivní semenné rostliny předcházející dnešním jehličnanům, které byly zastoupeny především kordiaty (Cordiatidea). Kordiaty měly hladký zhruba 20 m vysoký kmen s bohatě rozvětvenou korunou a velkými pentlicovitými listy. Udrželi se do permu a koncem prvohor vyhynuly.
|
|
|
Obr. 7.5.6 Pecopteris plumosa, svrchní karbon, Lubná u Rakovníku. |

Počátkem pennsylvanu došlo k ochlazení, což vedlo ke změnám v jednotlivých klimatických pásmech ve složení flóry mezi tropickým a chladným pásmem. Chladnější oblasti byly bohaté na semenné typy rostlin – hlavně Pteridospermy. Chladné oblasti jižní polokoule charakterizují rody Glossopteris a Gangamopteris.
Mořská fauna mississippu navazuje na vývoj devonského společenstva, ovšem mění se některá rodová složení a zastoupení čeledí. Během karbonu vyhynuly pancéřnaté ryby, tabulátní koráli, stromatoporoidi, nautilodní hlavonožci i trilobiti.
|
|
|
|
Obr. 7.5.7 Goniatites sp. svrchní karbon, Karviná. |
Obr. 7.5.8 Goniatites striatus, spodní karbon, Kyjovice. |


|
|
|
Obr. 7.5.9 Lingula squamiformis, spodní karbon, Ostrava. |

Naopak velkého rozmachu dosáhli mořští prvoci ze skupiny dírkovců (Foraminifera). Některé rody se vyskytovaly ve značné míře a měly významný horninotvorný význam. Maximálního rozvoje dosáhly lilijice, hlavonožci a zvláště goniatiti sloužící ke srovnávání mořských vrstev.
Karbon je charakteristický výrazným rozvojem suchozemských členovců, především čeledí hmyzu, stonožek, štírů, pavouků atd. Nejčetnější byl i tehdy hmyz. Tomuto rozmachu odpovídá příznivé klima a potrava ve formě nespočetné flóry. Střední karbon byl dobou hmyzu, tehdejší kontinent byl hmyzem přímo zaplaven. Mezi karbonskou dominantu patří obrovská dravá pravážka Meganeura, která s rozpětím křídel 75 cm patřila mezi největší druh hmyzu. Žilo spoustu dalších hmyzích řádů, které ke konci prvohor vymřely. Zajímavou vymřelou hmyzí skupinou byli pramřížonerví (Palaeodictyoptera) se šesti křídly. Zvláštním a současně velkým karbonským členovcem obývajícím karbonské pralesy byla zhruba 150 cm dlouhá stonožka Arthropleura. Její fosilní pozůstatky se vzácně našli i na našem území.
Vodní prostřední bylo plné života. Ve vodách byli hojní červi, mlži, břichonožci, drobní korýši a výjimečně i hrotnatci (Merostomata) – předchůdci ostrorepů. Sladké karbonské vody obývali drobní akantodi, ryby kostnaté (dvojdyšné, paprskoploutvé) a také velcí pražraloci z řádu Xenacanthida s typickým mohutným trnem, který jim vyrůstal těsně za hlavou. Vlhké břehy jezer, močály i řeky obývali krytolebci. Krytolebci vyvinutí z lalokoploutvých ryb neměli na souši žádnou konkurenci, a tak se hojně rozmnožovali. Tito obojživelníci jsou charakterističtí pro karbon a spodní perm, a proto se tato období mladších prvohor často označují za dobu obojživelníků. Krytolebci mohli dorůstat do velikosti jen pár cm anebo až do velikosti dnešních krokodýlů. Někteří se podobali spíše ještěrkám, jiní mlokům či hadům. Tělo měli pokryto pancířem z jemných šupinek a lebka byla zcela pokryta kostmi, odtud pochází jejich pojmenování. Do vody snášeli vajíčka, ze kterých se líhly larvy dýchající vnějšími žábrami.
Koncem karbonu, začátkem permu přišla změna. Vlhké a teplé podnebí vystřídalo období sucha. Ubývá vlhkých pralesů a přibývá suchých pustin. Tyto změny měly za následek vymizení mnoha rodů karbonské flóry, ale také měly vliv na vývoj fauny. Živočichové závislí na vodě se museli stáhnout do oblastí, kde voda ještě stále přetrvávala. Krytolebci se sušším podmínkám dokázali přizpůsobit. Jejich těla pokryla zrohovatělá kůže chránící organismus před rychlým vypařováním vody. Vajíčka již nesnášeli do vody, ale do tlejícího rostlinstva a písku. Vodu potřebnou k vývoji zárodků vystřídala tekutina v amnionové dutině vajíček (mají dnešní ptáci, plazi, savci a obratlovci ze skupiny Amniota). Změnila se také stavba kostry. Vznikla významná skupina, první plazi. Důležitý mezník v historii - došlo k úplnému přechodu na souš. První plazi se objevili ve svrchním karbonu, ale větší rozvoj je čekal až v permu.
Ve spodním karbonu v Evropě, který navazuje na mořský devonský režim, můžeme rozlišovat dva hlavní mořské vývoje: kulm a uhelný vápenec. Kulmský vývoj je charakteristický klastickými sedimenty, tj. střídáním drob, resp. slepenců s prachovci a jílovitými břidličnatými sedimenty. Uloženiny mají flyšový charakter, fauna bývá chudá, ichnofosilie patří hlubokovodní (nereitové) ichnofacii.
|
|
|
Obr. 7.5.10 Sedimenty spodního karbonu – kulm, Stará Ves. |

Vývoj uhelného vápence je charakteristický mělkovodní karbonátovou sedimentací v teplém klimatickém pásmu. Bývá označován jako dinant. Uloženinami jsou bioklastické karbonáty, většinou s hojným bentosem.
|
|
|
Obr. 7.5.11 Uhlonosný svrchní karbon paralické pánve, vrch Landek. |

Nejtypičtějším sedimentem karbonu je černé uhlí. Výchozí materiál vznikal za vlhkého klimatu v prostředí močálů s bujně rostoucí vegetací. Podmínky pro uhlotvornou sedimentaci byly příznivé zejména v tektonicky podmíněných, intenzívně klesajících mezihorských svrchnokarbonských pánvích. Uhlotvorná prostředí vznikala v pánvích dvojího typu. V pánvích limnických, ve sladkovodních a kontinentálních podmínkách, v depresích a mezi hřbety vyvrásněných pohoří a v pánvích paralických v přímořských oblastech, ve kterých se střídal sladkovodní a bažinný režim s občasnými mořskými záplavami. Uhlonosné pánve prvého typu se vyvíjely především ve vyšším svrchním karbonu. Jejich sedimentace obvykle pokračovala více-méně bez přerušení dále do permu, takže jejich uloženiny bývají spojovány s termínem permokarbon. Sedimentace v paralických pánvích bývá ukončená variským vrásněním před koncem karbonu ve stupni westphal.
|
|
|
Obr. 7.5.12 uhlonosný svrchní karbon limnické pánve, Žacléř. |

Karbonský útvar byl dobou významných horotvorných pochodů, které shrnujeme pod termín variské (hercynské) vrásnění. Variské horotvorné procesy byly v několika fázích provázené pohyby a deformacemi velkých horninových celků, zlomy, metamorfózou, tavením, plutonickou a vulkanickou činností, ale též erozí a obnažováním těles nacházejících se dříve v hloubce. Postupně se formující variské pásemné pohoří vytváří oproti spodnímu karbonu zcela jiné sedimentační podmínky. Jsou spojené se vznikem rozsáhlých hornatých souší, ale též s přímořskými plošinami, depresemi a jezerními pánvemi, zarůstanými bujnou karbonskou vegetací, často za doprovodu vulkanismu.
Variské horstvo lze podle koncepce F. Kossmata rozdělit do několika podélných pásem západo-východního směru s různou intenzitou a stářím deformací. Nejhlouběji obnažená, nejsilnější metamorfózou postižená, variskými granitoidními masivy prostoupená nejvnitřnější zóna pohoří se označuje jako moldanubikum. Variské projevy lze zde sledovat od vyššího devonu do spodního visé.
Severněji ležící pásmo se označuje jako saxothurigikum. Je rovněž postižené regionální metamorfozou. K hlavním procesům vrásnění došlo většinou během nebo až na konci spodního karbonu (sudetská fáze). Intruze plutonitů, ale též výlevný suchozemský vulkanismus, proběhly v karbonu až v permu. Postorogenní stadium je spjaté s rozsáhlou permokarbonskou uhlotvornou sedimentací.
Moldanubikum a saxothurigikum náležejí k variským internidám. Na jejich obvodu vystupuje na povrch velmi úzké pásmo se silně metamorfovaným prekambriem a starším paleozoikem. Je označované jako severoněmecký práh.
Zóna variských externid se k internidám přimyká pásmem rhenohercynika. Je to pásmo, které se vyznačuje intenzivní devonskou a karbonskou subsidencí a odpovídající sedimentací. Zóna subsidence se postupně stěhuje (migruje) od nitra variscid ve směru do předpolí. Devon začíná transgresivně. Devonské a spodnokarbonské uloženiny byly postupně zvrásněny v období svrchní visé až westphal (asturská fáze) a sedimentace ukončena. Variský plutonismus je v tomto pásmu vzácný.
Subvariscikum představuje nejsevernější pásmo variscid, které se vytvořilo na konsolidovaných částech kratonů v předpolí variscid. Pásmo tvoří většinou uhlonosné svrchnokarbonské sedimenty, usazené ve vnějších molasových pánvích. Variské vrásnění zde doznívalo až v pozdním karbonu, takže uloženiny nejmladšího karbonu, pokud se vyvinuly, jsou již nezvrásněné.
7.6
Perm (299,0 +-0,8 až 251,0 +- 0,7 miliónů let)
Název perm je odvozen od města Perm na západním Uralu. Jako samostatný útvar byl vymezen Murchisonem v r. 1841.
Rozdělení permského útvaru na jednotky prodělalo v posledních létech dosti podstatné změny, když se ve vhodných mořských uloženinách podařilo schválit mezinárodně platné stupně v mořském vývoji, stejně tak jako definovat spodní a svrchní hranici permu. Značné rozšíření souší v permu v důsledku doznívajícího variského vrásnění je spojeno s převážně kontinentální sedimentací, ve kterých už mnohem dříve byly stanoveny kontinentální stratigrafické jednotky. Ty ovšem mají pouze regionální platnost.
Hranice mezi karbonem a permem je stanovena na základě prvního výskytu konodonta Streptognathodus ,,wabaunsensis“, tento stratotyp se nachází u Aidaralašského potoka na jižním Uralu v severním Kazachstánu. V kontinentálním prostředí hranice karbonu a permu je vypozorována nápadným úbytkem výtrusných rostlin, a naopak rozvojem rostlin nahosemenných (zejména pteridosperm). Nejmladší útvar paleozoika – perm má spodní hranici datovanou na 299 milióny let a svrchní hranici na 251 milióny let. Permský útvar se nově dělí na mezinárodně schválená tři oddělení a devět stratigrafických stupňů. Perm se dělí do oddělení cisural, guadelup a lopping. Ve střední a západní Evropě se perm v kontinentálním vývoji dělí tradičně na dvě oddělení – na perm spodní (v německé literatuře též červená jalovina) a svrchní (též zechstein).
Podnebí v permu navazovalo na svrchní karbon, stávalo se stále sušším a počátkem guadelupu i značně teplejším. Ledovec pokrývající mocná území na jižní Gondwaně značně ustoupil, čímž byla ukončena téměř 50 miliónu let dlouhá doba ledová. Roztávání ledovce vedlo k celosvětové transgresi.
Pangea byla před přínosem srážek do vnitrozemí chráněna horskými hřebeny, a tak zde vznikaly rozsáhlé pustiny s charakterem polopouští až pouští typické pro převážnou část oblastí Evropy a Severní Ameriky. Kontinentální sedimenty permu této oblasti jsou nejčastěji červeně zbarvené.
Uhlotvorná sedimentace pokračující z karbonu až do středního, místy až svrchního permu se přesunula z oblastí okolo rovníku do vyšších zeměpisných šířek, kde přetrvávalo příznivější srážkové podnebí.
|
|
|
Obr. 7.6.1 Mělkomořské pelitické sedimenty doprovázené bahenními prasklinami a evapority, Sczavnica, Polsko. |

|
|
|
Obr. 7.6.2 Arkózové sedimenty permu se silicifikovaným kmenem, dolnoslezská pánev, Žaltman. |

|
|
|
Obr. 7.6.3 Bahenní praskliny patří k běžným znakům sedimentů svrchního permu. |

Během permu došlo ke srážce Siberie a Kazachstánie s Laurusií, v místě srážky se vyvrásnilo pohoří Ural. Spojení samostatných kontinentů vedlo ke vzniku superkontinentu Pangea. Některé menší kontinenty si udržely svou samostatnost až do počátku mezozoika.
Doznívaly hercynské horotvorné pochody. Doznívání variských horotvorných procesů provází často intenzivní vulkanická činnost.
|
|
|
Obr. 7.6.4 Poloha kontinentů ve svrchním permu (šipky označují směr trangrese v zechsteinu na Laurussii). |
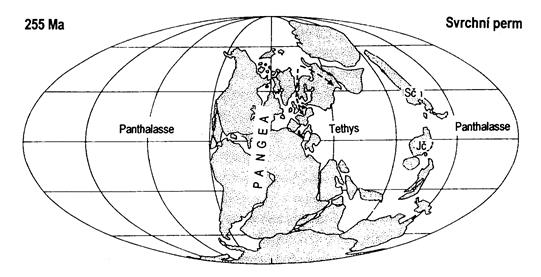
Život rychlým tempem pokračoval ve svém vývoji jak ve vodě, tak na souši i ve vzduchu. Ve spodním permu přetrvávají výtrusné rostliny, hlavně kapradiny (Callipteris, Pecopteris, Ondotoptersi), plavuně (Sigillaria) a přesličky (Calamites), ale ve svrchním permu dominují již rostliny nahosemenné. Ve spodním permu se začínají objevovat jinanovité a cykasovité rostliny a především pak jehličnany, kterým vyhovovalo sušší podnebí.
Během svrchního permu dosáhlo rostlinstvo vyššího vývojového stupně, na rozdíl od vývoje živočišného, který byl poněkud zpomalen.
|
|
|
Obr. 7.6.5 Walchia sp., spodní perm, dolnoslezská pánev (sbírka VŠB-TU Ostrava). |

Mořská fauna ve spodní části permu navazovala na karbon, avšak hned počátkem svrchního permu docházelo k vymírání, které gradovalo ke konci permu. Toto vymírání bylo nezřetelnější v tropických mělkovodních společenstvech pravděpodobně následky mocných výlevů bazaltů v oblasti Sibiře, kdy při tuhnutí se do atmosféry dostalo velké množství oxidu uhličitého.
Dírkovci, čtyřčetní koráli a ramenonožci, mechovky, goniatiti, trilobiti koncem permu vymřeli. Teplé klima prospívalo k tvorbě organických útesů tvořených především vápnitými řasami, houbami, mechovkami či lilijicemi.
Některé skupiny obratlovců např. ryby a obojživelníci jsou také na ústupu. Vývoj u obratlovců pocítili hlavně plazi. V tomto období se běžně objevuje mnoho čeledí krytolebců, kteří z velké části během svrchním permů vymírají a triasu se dožívají už jen ti nejsilnější.
|
|
|
Obr. 7.6.6 Zástupce krytolebců Discosauriscus sp., Obora. |

Změna klimatu, která měla za následek vznik rozsáhlých vyprahlých oblastí, způsobila ústup obojživelníků. Na druhé straně však nastavila podmínky ideální pro rozvoj plazů. Za krátkou dobu vznikly četné formy plazů z řádů Captorhinomorpha a Pelycosauria. Řád Captorhinomorpha představuje mohutné plazy značně připomínající kryptolebce. Tato nejprimitivnější skupina plazů byla masožravá i býložravá. Řád Captorhinomorpha se rozčlenil do dvou skupin, kdy u jedné vývoj pokračoval přes různé typy plazů k savcům a následně až k člověku. Druhou skupinu reprezentují předkové dnešních ještěrek, želv, nebo např. tříoké haterie žijící na Novém Zélandu. Druhý řád Pelycosauria – pelykosauři byli primitivní a studenokrevní plazi. Teprve ke konci permu se objevuje vyspělejší větev již teplokrevných savcovitých plazů (Therapsida). Tito plazi žili v chladnějších klimatických podmínkách, a proto jejich těla byla pokryta srstí.
Obdobím permu skončila éra prvohor trvající zhruba 345 miliónů let. Život se z moře pozvolna přesunul na souš, kde panovalo sucho a ne příliš pohostinné podmínky. Nově vytvořené formy se těmto podmínkám přizpůsobily a dokonce z nich vznikly linie pozvolna směřující k vývoji savců.
Na konci permu je nápadné hromadné vymírání bezobratlých živočichů. Bývá spojováno s katastrofickou událostí, která ale není jednoznačně doložená. V kontinentálních prostředích plazi zatlačují do pozadí obojživelníky (krytolebce). K podstatným změnám též dochází ve složení suchozemského rostlinstva. Nahosemenné rostliny ve vyšším permu začínají převažovat nad dosud dominujícími rostlinami výtrusnými (nástup mezofytika).