8. Mezozoikum
Mezozoikum
představuje druhou velkou éru historie Země, trvající zhruba 160 miliónů let.
Mezozoikum započalo přibližně po zakončení hercynských horotvorných pochodů a
ukončeno bylo během jedné z rozsáhlých regresí na hranici křídy a
paleocénu. Hranice také určuje evoluce
ba naopak vymření daných živočišných skupin.
Rozsáhlá pevnina Pangea sdružující kontinenty
počátkem mezozoika se postupem času začíná rozpadat a formovat rozsáhlé mobilní
zóny v oblasti Tethydy,
Atlantského, Indického a Severního ledového oceánu. Během
mezozoika proběhly hlavní fáze alpinské orogeneze, které vedly k uzavírání
oceánu Tethys. Alpinská orogeneze se dělí na tzv. staroalpínskou fázi probíhající především
v křídě a mladoalpínskou probíhající v terciéru.
Řetěz mladých pásemných pohoří s typickou příkrovovou stavbou
v dnešní době začíná v severní Africe Atlasem, pokračuje přes Betickou kordileru do Pyrenejí,
Alp, Karpat, Apenin, Balkánu na Kavkaz a dále pokračuje do Asie do Himalájí.
Mezozoikum se dělí na tři útvary označované
jako jura, trias a křída. Život v mezozoiku byl zcela
jiný než život v paleozoiku, fauna i flóra se značně změnila. Vůdčí postavení
v bentózních společenstvech během paleozoika
ztrácejí brachiopodi, které biologicky nahrazují plži a především mlži. Hranici mezi paleozoikem a mezozoikem nepřekročila řada
živočichů typické paleozoické fauny (např. trilobiti, graptoliti,
rugózní a tabulátní koráli,
cystoidní a blastoidní
ostnokožci, fusulinidní dírkovci). Ve velkém byly v
moři redukovány mechovky, ramenonožci a na souši obojživelníci. Naopak se
objevují nové skupiny živočichů, jako jsou amoniti
se složitým amonitovým typem švu a belemniti.
Amoniti patřili k nejběžnějším fosiliím mořského prostředí, kdežto souš
ovládli dinosauři, a to mnoho forem
býložravých, všežravých i masožravých. Vodní prostředí ovládli vodní ještěři a ve vzduchu se proháněli
ptakoještěři. Počátkem mezozoika se
objevují první savci i první ptáci. Ve flóře měly hlavní
převahu nahosemenné rostliny. Na
konci křídy došlo k diverzifikaci krytosemenných rostlin. Obdobně jako na
hranici paleozoika a mezozoika, tak i hranici mezozoika a kenozoika byla
zakončena velkým vymíráním.
|
|
|
Obr. 8.1 Vymezení mezozoika. |
8.1 Trias (251,0 ±
0,7 až 199,6 ± 0,6 miliónů let)
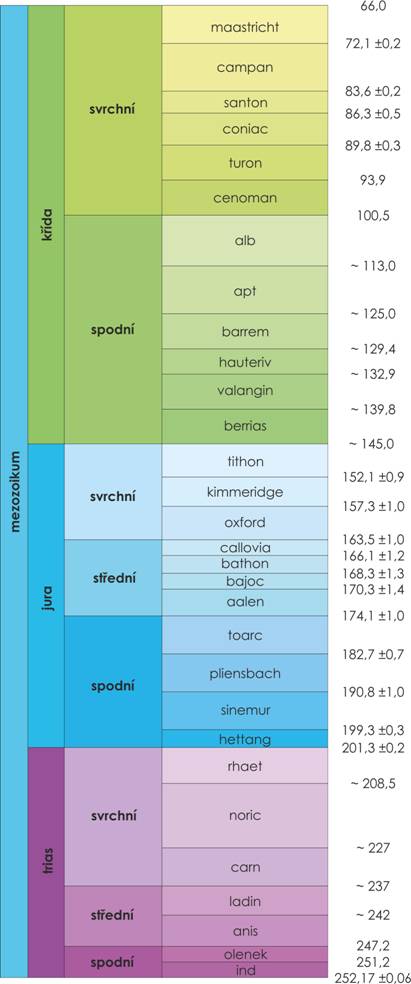
Triasový útvar vyčlenil v roce 1934 němec August von Alberti. Název trias je odvozen od tří sedimentárních celků v typickém evropském vývoji. Nejstarší pískovce na bázi jsou označovány jako Buntsandstein – pestrý jílovec, střední jako Muschelkalk – lasturnatý vápenec a v nadloží nejmladší jako Keuper – pestré slíny a břidlice. Stratotyp spodní hranice triasu je stanoven na základě prvního výskytu konodonta Hindeodus parvus v oblasti východní Číny. Stratotyp svrchní hranice nebyl dosud přesně biostratigraficky vymezen. Trias se dělí do tří oddělení, podní, střední a svrchní. Oddělení spodního triasu se dělí na stupně ind a olenek. Oddělení středního triasu má stupně anis, ladin a oddělení svrchního triasu má stupně carn, noric a rhaet.
V Evropě rozeznáváme dvojí typ vývoje sedimentace, jedná se o vývoj germánský a alpínský. Germánský vývoj triasu tvoří sedimenty pevninské, mořské, lagunární i brakické. Vrstvy mocné jen několik set metrů se usazovaly v oblastech střídavě zaplavovanými mořem. Tyto oblasti byly v době mezi záplavami zarostlé hojným rostlinstvem. Trias alpínského vývoje sedimentoval v mořském prostředí. Usazovaly se mocné vrstvy vápenců a dolomitů, a jelikož se ukládaly v klesajících pánvích, dosáhly větších mocností.
Od tethydního sedimentačního prostoru, kde se ukládají uloženiny alpinského vývoje triasu, je epikontinentální (germánský) vývoj oddělen tzv. vindelickou pevninou. Jejím jádrem, ale též severovýchodním ukončením je Český masív.
|
|
|
Obr. 8.1.1 Paleogeografická rekonstrukce
území střední Evropy během středního triasu. |
U germánského i alpského vývoje přechází perm
většinou bez znatelných hranic do spodního triasu, stejně tak je to i u svrchní
hranice mezi triasem a jurou. Podnebí v triasu bylo vyrovnanější než
v permu. Bylo velmi teplé i velmi suché. Důkazy o zalednění polárních
oblastí chybí, naopak byly tyto oblasti obydleny plazy. Na hranici permu a
triasu došlo ke značnému snížení hladiny světového oceánu. Do středního triasu
probíhalo střídavé zvyšování mořské
hladiny. Během středního triasu proběhla výrazná regrese a opětovné
zvyšování hladiny oceánu. Hranici triasu a jury vystihuje znovu výrazná
regrese. Sedimenty triasu jsou výsledkem suchého
a aridního vývoje, čemuž nasvědčují krusty, evapority, ložiska solí a
sádrovce.
|
|
|
Obr. 8.1.2 Superkontinent
Pangea ve svrchním triasu. Šipky označují komunikaci
epikontinentálních moří v Evropě (podle Faupla,
2000). |
Pro období
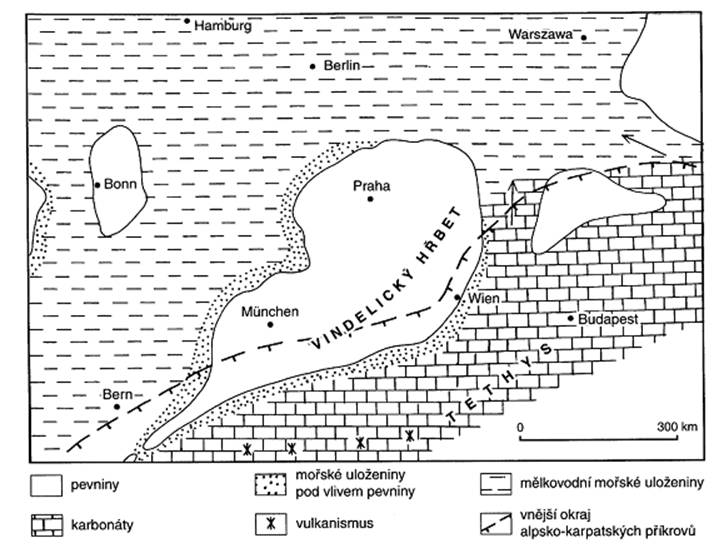
trias je typický jednotný superkontinent Pangea obklopený oceánem Panthalassa. Na
východní polokouli se rozprostíral oceán Paleontethys. Ve svrchním triasu
byly oživeny riftové struktury, které vedly k posunu mikrokontinentů
ve směru k eurasijskému mobilnímu prostoru. Tyto pohyby zapříčinily
uzavření oceánu Paleotethys a následnému otevření
nového oceánského prostoru Neotethys. Kolize těchto mikrokontinentů
s euroasijskou pevninou je označována jako starokimerská fáze. Prostor oceánu Tethys se v triasu postupně klínovitě rozšiřuje
k západu, což naznačuje budoucí rozdělení Pangey na severní a jižní
skupinu kontinentů. Na jihovýchodě se začíná zakládat prostor budoucího
Indického oceánu.
Triasové
klima se svou ariditou podobalo permskému. Široký
suchý pás pod a nad rovníkem, charakteristický sedimentací červených a pestrých
sedimentů, zatlačil pásma vlhkého (humidního) klimatu dále směrem k jihu a
severu, kde pod vlivem monzunových dešťů byly podmínky vhodné pro uhlotvornou vegetaci (dnešní severovýchodní Asie a jižní
část Gondwany).
|
|
|
|
Obr. 8.1.3 Eolické sedimenty spodního
triasu, Rtyně v Podkrkonoší. |
Obr. 8.1.4 Pestré sedimenty svrchního
triasu s pozůstatky plazů a obojživelníků, Krasiejow,
Polsko. |


|
|
|
Obr. 8.1.5 Uhlonosné sedimenty triasu,
Rumunsko. |

V tropické klimatické zóně Tethydy se v triasu na rozlehlých karbonátových
platformách ukládaly mocné soubory karbonátů různého typu (vápence, ale též
primární dolomity). Dnes jsou známé především z oblasti Severních
Vápencových Alp, Dolomit, Západních Karpat aj.
|
|
|
Obr. 8.1.6 Sedimentární dolomit, Dolomity,
Itálie. |

Na hranici permu/triasu postihlo
živočišnou existenci největší světové vymírání.
U mořských druhů se vegetace snižuje až o 95%, na kontinentech až o 70%. Vymírání
proběhlo velmi rychle a mělo katastrofické následky, které postihly některé
skupiny živočichů v jejich maximálním rozvoji, naopak rostlinstvo
nezaznamenalo žádné větší změny. Pravděpodobně za vymírání mohl prudký pokles
hladiny světového oceánu spojený s úbytkem kyslíku v mořské vodě
(tzv. anoxie). Důsledkem poklesu hladiny oceánu se zmenšily oblasti
kontinentálních šelfů. Příčiny
vymírání jsou také spojeny s klimatickými změnami vyvolanými vulkanickou
aktivitou na střední a západní Sibiři. Do atmosféry se uvolnilo velké množství
hydrátu metanu a snížil se obsah kyslíku na 13%. Tento ekologický stress přežily pouze životaschopné skupiny živočichů.
Od svrchního permu až do počátku svrchní
křídy převládaly nahosemenné rostliny,
které se vývojem přizpůsobily suššímu prostředí. Hojní byli zástupci cykasovitých rostlin. Velký význam měla vymřelá skupina Bennettitales (rostliny benetitové) křovitého či
stromovitého typu. Z paleozoické flóry se stále ještě držely obrovské
stromovité přesličky a plavuně čeledi Gleicheniaceae. V rozvoji pokračují jehličnany, cykasy
a ginkgovité rostliny.
|
|
|
Obr. 8.1.7 List jinanu, Rumunsko. |
Triasové moře se lišilo od moře permského. Čtyřčetné a deskaté korály
nahradili koráli šestičetní, kteří
byli tvořeni šesti přepážkami stejné velikosti. Rozšířily se některé skupiny ramenonožců, ostnokožců, pravidelných
ježovek, mlžů, břichonožců.
K převládajícím obyvatelům moří v triasu zaručeně patřili hlavonožci, přežívaly i starší typy loděnkovitých, ovšem maximálního rozvoje dosáhli amoniti. V tomto období vládli ceratiti, phyloceratiti, lytoceratiti se složitým švem. Rozmohli se také hlavonožci dvoužábří, ke kterým patří belemniti. Během triasu vymírají konodonti i konulárie.
Triasové útesy vytvářeli krinoidi, spongie, vápnité řasy, dále v menší míře mlži a
ramenonožci.
|
|
|
Obr. 8.1.8 Ceratites nodosus, střední trias, Německo (foto P. Skupien). |
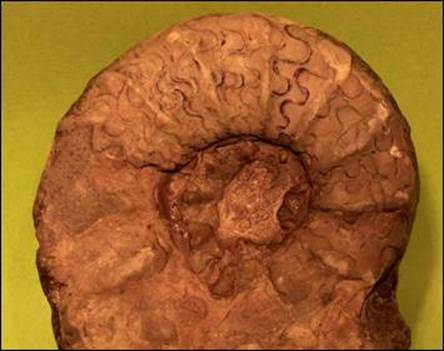
Některé ryby přežily až do počátku mezozoika. V mořích bylo možné
vidět některé druhy lalokoploutvých ryb,
dvojdyšných ryb a paprskoploutvých ryb.
Krytolebci ještě stále hojně obývali triaské
souše, ke konci triasu však vymírají. Nastal obrovský rozvoj plazů, kterým patřil tehdejší druhohorní svět.
|
|
|
|
Obr. 8.1.9 Metoposaurus sp., svrchní trias, Polsko. |
|


Na pevnině se objevují žáby, které představují evoluční
novinku. Obojživelníci v tuto dobu dorůstali největších velikostí
v celé historii Země. Objevují se létající plazi – pterosauři. V triasu dále postupoval rozvoj savcovitých plazů. Savcovití
plazi jsou velice důležití z vývojového hlediska. Z primitivní permokarbonské skupiny (Pelycosaria) se vyvinuly koncem
permu dvě skupiny, a to masožraví theriodonti (Theriodontia) a
převážně býložraví anomodonti (Anomodontia). Z masožravé
skupiny theriodontů vznikli během triasu první savci.
Ze skupiny archosaurů (Archosauria) se oddělily linie
vedoucí ke krokodýlům a k dinosaurům.
Na začátku mezozoika,
v triasu se objevují první zástupci skupiny, která ovládne následujících
180 milionů let. Skupinu Dinosauria v překladu
,,hrozní plazi“ pojmenoval Angličan Richard Owen. Velmi brzy se
rozdělili na dvě nejpodstatnější linie, řád plazopánví
(Saurischia)
a ptakopánví (Ornithischia). Ptakopánvé
zastupovali býložravci, plazopánvé jak býložravci,
tak i masožravci. Na konci triasu se
objevují již velké formy dinosaurů dosahující délky až 12 m.
Během
svrchního triasu proběhlo další velké vymírání, které postihlo především
suchozemské ekosystémy. Od vymírání, které probíhalo na hranici perm/trias je
hlavní rozdíl v tom, že mořské ekosystémy nebyly téměř dotčeny. Na souši
vymřelo poměrně v krátko době zhruba 25% druhů, především vývojové linie
plazů (teromorfní plazy). Důvody nejsou zcela známy.
Starší hypotézou je pád meteoritu, ovšem nové výzkumy poukazují na vulkanickou
činnost, která sílila počátkem rozpadu Pangey.
8.2
Jura (199,6 ± 0,6 až 145,5 ± 4,0 miliónů let)
Název
jurského útvaru je odvozen od pohoří Jura, které
leží v předpolí Alp ve Švýcarsku a Francii. Pojmenování zavedl již na konci 18.
století vynikající německý přírodovědec Alexandr von Humboldt.
Spodní hranice jury
v mořském prostředí je dána výrazným vymíráním zejména amonitové fauny na
konci triasu a je prokázána výraznou změnou v jejím složení. Juru dělíme
na oddělení spodní jury – lias,
střední jury – dogger a svrchní jury
– malm. Jura se dělí na jedenáct stratigrafických
stupňů. Oficiálně schválený stratotyp spodní hranice
má však jen stupeň bajoc (lokalita Murtinheira v Portugalsku). Jurský útvar představuje
jediný sedimentační cyklus, který byl započat transgresí a ukončen regresí.
Pro stanovení spodní a svrchní
hranice jurského útvaru, stejně jako pro bližší členění na stupně mají rozhodující význam amoniti. Spodní
hranici indikují prvé výskyty amonita Psiloceras planorbis. Široce diskutovaným problémem zůstávají
vztahy mezi studenovodními (boreálními)
a teplovodními (tethydními)
faunami.
Podnebí
v juře bylo stabilní a velmi teplé. Ve spodní juře se výrazně oteplilo,
což se v mořích projevilo anoxií
(doloženo výskytem tmavých bituminózních břidlic). Ve spodní juře se šířila mořská
transgrese, které zalila rozsáhlé oblasti Evropy a Asie. Moře proniklo na území
Českého masívu do severních Čech a na Moravu, dále do horního Slezska až
Polska. Moře bylo mělké s převládající karbonátovou sedimentací.
Ve svrchní juře probíhalo nové vrásnění,
v Evropě označované jako mladokimerské a v Americe jako nevadské. Vrásnění dalo
za vznik novým pásemným pohořím, které byly doformovány ve svrchní křídě.
Kontinent Gondwana byl již rozčleněn na jednotlivé
světadíly. Ve svrchní juře se začala odtrhovat Indie od Antarktidy a
vznikly riftové struktury mezi Antarktidou,
Jižní Amerikou, Austrálií a Afrikou.
Stopy zalednění v juře nejsou známy. Teplomilná flóra rostla i ve vyšších
zeměpisných šířkách. Podnebí bylo pravděpodobně teplejší jak dnes, ale ne tak
suché jak v triasu. Ve svrchní juře vznikla znovu podnebná pásma.
|
|
|
Obr. 8.2.1 Uspořádání kontinentů před 160 Ma (kolem hranice dogger/malm). |
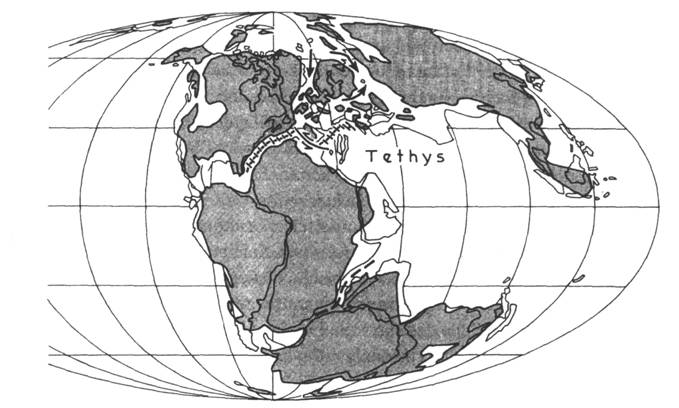
Paleogegografické změny měly za následek nárůstání ploch šelfů v teplém klimatickém pásmu. To přispívalo k rozvoji mělkovodních mořských faun. Tento obraz je v souladu s vysokým stavem oceánské hladiny. Vrcholu je dosaženo v hraničním intervalu střední a svrchní jury, ve stupních callov a oxford. Mělkým mořem je tehdy zalita většina západní Evropy a západní části Severní Ameriky. Tato událost je známá pod názvem callov- oxfordská záplava. V uvedeném období mimo jiné též došlo k vytvoření mělkého mořského průlivu přes Český masiv (ve směru SZ – JV) a rovněž k jurské transgresi přes jihovýchodní okraj Českého masivu.
Jurská flóra je zastoupena cykasy, benetity, ginkgovitými rostlinami, jehličnany, kaytóniemi a bylinnými kapradinami. V juře se naposledy v geologické historii Země objevovaly stromovité přesličky (rod Neocalamites). V teplém tropickém prostředí se dařilo stromovitým kapradinám, v mírnějších podnebných pásmech spíše nižším, bylinným rostlinám. Dominantními rostlinami jury byly rostliny nahosemenné, k nejnápadnějším patřily cykasy. Vyhynulá skupina rostlin benetitových (Bennettitales), značně rozšířená v juře byla pravděpodobně předchůdcem rostlin krytosemenných. Běžné byly rostliny jehličnaté (předchůdci dnešních borovic, cedrů, sekvojí i cypřišů). Dalšími četnými skupinami byly rostliny blahočetovité (Araucariaceae) a tisovcovité (Taxodiaceae). V mořském prostředí se rozvinuly mořské řasy, typický ruduchovitý rod Solenopora a ze zelených řas běžné čeledi Dasycladaceae.
Ve svrchním triasu živočišný svět ovládli dinosauři, kteří jsou známí ze všech kontinentů. Dinosauři koncem jury dosáhli maximálních velikostí. Představovali největší suchozemské predátory a býložraví dinosauři největší suchozemská zvířata všech dob. Početný hmyz přilákal dinosaury do vzduchu, kde konkurovali pterosaurům, kteří byli doposud dominantní letci jurského období. Savci v období jury nedosahovali větších velikostí a pravděpodobně žili nočním životem. Většina vývojových linií savců neměla dlouhé trvání. Během jury vymřeli poslední terapsidní (teromorfní) plazi.
V mořích se opětovně rozvinuli prvoci ze skupiny dírkovců. Velký rozvoj poznamenal i šestičetné koráli, společenstva měkkýšů, křemitých hub a mechovek. Významným jurským nalezištěm je Kotouč u Štramberku. Ramenonožcům se nedařilo, jejich úbytek nahradili mlži. Nejdůležitějšími hlavonožci jurského moře byli amoniti. Mezi běžné obyvatele moří patřili kostnaté ryby, žraloci, rejnoci. Moře ovládli vodní plazi, obzvláště ryboještěři, pleistosauři. V mělkých mořích se dařilo lilijicím, ježovkám, hadicím, krabům a jiným korýšům.
|
|
|
Obr. 8.2.2 Sediment střední jury tvořený
křemitými houbami a amonity, Zalas Polsko. |

|
|
|
|
Obr. 8.2.3 Hildoceras
bifrons, spodní jura, Ladce,
Slovensko |
Obr. 8.2.4 Quenstedtoceras
henrici, spodní jura, Polsko. |


|
|
|
|
Obr. 8.2.5 Krinoidový vápenec s rostry
belemnitů, střední jura, Ladce, Slovensko. |
Obr. 8.2.6 Vápenec s amonity rodu Hildoceras sp., Ladce, Slovensko. |


|
|
|
|
Obr. 8.2.7
Zástupci svrchnojurských korálů, Štramberk, Stylina tubulifer. |
Obr. 8.2.8 Montlivaltia cf. rosula. |
|
|
|
|
Obr. 8.2.9 Stylina ostravensis. |
Obr. 8.2.10 Latomeandra fromenteli. |




|
|
|
|
Obr. 8.2.11 Heterodiceras
lucii, Štramberk, svrchní jura. |
|


Součástí sladké a slané jurské
vody byli praví krokodýli (Crocodilia)
vyspělí z jamkozubých předků během triasu.
Podtřída Archocauria přežila věk plazů a dožila se dnešní
doby. Dnes obývají krokodýli sladké vody, ale v juře žila většina
v mořích.
Dinosauři se vyvinuli skoro
současně s krokodýly a ptakoještěry v období triasu
z primitivních jamkozubých předků. Koncem triasu
se rozšířily některé skupiny dinosaurů a do konce druhohor ovládli Zemi.
Jednotlivé druhy dinosaurů bylo velice odlišné, někteří dosahovali velkých
rozměrů a hmotnosti několik tun, další velikosti maximálně 25 cm. Všichni však
měli velikou výhodu, a to velmi rychle se pohybovat po souši.
Dinosauři čeledi plazopánví
(Saurischia)
měli těla velmi podobné jamkozubým předchůdcům. Měli
dlouhý a dobře vyvinutý ocas, který u dvounohých forem (bipedních) sloužil jako
balanční orgán. Ocas se také zachoval u forem kvadrupedních,
to jsou formy chodících po čtyřech. Řád Theropoda představuje
masožravé dvounohé formy, ovšem v průběhu evoluce některé rody přešly
z dvounohého způsobu života na kvadrupední
živící se rostlinou formou, z kterých se vyvinuly obrovské formy
dinosaurů. Řád Saurichia
měl v poměru k tělu malou lebku a zuby po celé délce čelisti.
Masožravé formy měli poslední články prstů zahnuté s mohutnými drápy.
Dvounohé formy měli vyvinuté mohutné zadní nohy. Povrch těla byl pokryt
kulatými či čtvercovitými šupiny, které často vytvářely mozaiky. Vyskytovaly se
jedinci s kostěnými krunýři s různými výrůstky. Někteří zástupci těchto
dinosaurů mohli dosahovat délky až 31 m a hmotnosti 80 tun.
Druhá
skupina čeledi ptakopánvích (Ornithischia) byla od svých
počátků býložravá, včetně dvounohých forem. Čtvernozí dinosauři ze skupiny ptakopánvích měli na krunýři všelijaké výrůstky ve formě
rohů, štítu atd. Patřili mezi býložravce a měli většinou jen dva zadní zuby.
Bezzubá čelist byla ve tvaru rohového zobáku. Ještěři ze zmiňované skupiny
nedosahovaly takových velikostí jako skupina předešlá, ale měli neobvyklé
tvary. Neměli končetiny přizpůsobeny k dvounohému způsobu života jako masožravá
forma Theropoda.
Významnou a velmi
charakteristickou skupinu jurské fauny představovali ptakoještěři (Pterosauria).
Patřili mezi jediné plazy, kteří jako první z obratlovců ovládli vzdušné
prostředí. Ptakoještěrům se vyvinula létací blána původně sloužící ke zmírnění
pádu ze stromu, později k přeletu ze stromu na strom a ve finále
k aktivnímu letu. Ptakoještěři obývali pevniny, avšak jejich fosilní
pozůstatky byly objeveny v mořských sedimentech, proto lze předpokládat,
že žili jako dnešní ptáci rybáci. Jejich potravou byly ryby a hmyz, které
lovili v letu. Ptakoještěři se objevili během jury a vymizeli koncem
křídy.
V období jury se
objevují první praptáci. Velikostně
byli trošku větší jak dnešní holubi. Byli to špatní letci a obývali břehy jezer
a lagun. Nejnovější výzkumy ukazují, že předci praptáků představovali malí
jurští dinosauři z podřádu Theropoda. Zlomem ve vývoji byla přeměna plazích šupin
v peří. Přeměna šupin v peří souvisela nejspíš se zvýšeným oběhem krve.
Opeření praptáky chránilo jak proti chladu, tak i před slunečním zářením.
8.3
Křída (145,5 ± 4,0 až 65,5 ± 0,3 miliónů let)
Posledním nejdelším obdobím
mezozoika byla křída, která trvala zhruba 70 miliónů let. Křída svůj název
dostala podle psací křídy, která se
vyskytuje v platformním vývoji v nejvyšší části útvaru a je tvořená
převážně schránkami vápnitého nanoplanktonu ze skupiny kokolitek.
Z biostratigrafického hlediska v tethydní oblasti je jura datována nástupem amonitů druhu Berriasella jacobi odpovídající
zhruba bázi kalpionelové zóny Calpionella alpina. Svrchní hranice,
hranice s terciérem je velmi výrazná a spojena s hromadným vymíráním
mezozoické bioty. Stratotyp hranice křída/terciér se
nachází v El-Kef
v Tunisu. Křída se dělí do dvou oddělení,
do spodního a svrchního. Oddělení spodní křídy se dělí na stupně: berrias, valangin, hauteriv, barrem, apt a alb.
Oddělení svrchní křídy má stupně: cenoman, turon, coniac, santon, campan a maastricht.
Podnebí v křídě bylo proměnlivé. Ve spodní
křídě bylo podnebí teplé a vyrovnané. Ve svrchní křídě docházelo ke zvyšování
hladiny světového oceánu, k transgresím i regresím, ke zvětšování
vzdáleností mezi jednotlivými kontinenty, ke změnám hladiny CO2
v atmosféře, což vedlo ke klimatickým změnám. I přes výše zmíněné změny
bylo podnebí stále tak teplé, že v polárních oblastech nebyly ledovcové
čepičky (období ,,Green house“) a ve vysokých
zeměpisných šířkách běžně rostly opadavé jehličnany a žili dinosauři. Na
hranici spodní a svrchní křídy kulminoval skleníkový efekt. V oblasti
našeho území panovalo teplé, subtropické podnebí. Český masiv byl zaplaven
mělkým mořem.
Rozložení
kontinentů v křídě
nemělo ještě dnešní podobu. Jižní Amerika se oddělovala a pomalu
vzdalovala od Afriky, Afrika a Indie se vzdalovaly od Antarktidy, Severní
Amerika a Grónsko se vzdalovaly od Evropy. Vznikly nové zlomové struktury,
které vedly k oddělení Austrálie od Antarktidy ve svrchní křídě. Zmiňované
oddalování, přibližování, vznik zlomových struktur mělo za následek výzdvih
oceánských vulkanických hřbetů, které vytlačovaly vodní masy, a stoupající
hladina světového oceánu zaplavila mocná území kontinentů. Během svého maxima
záplavy pokryly celou jednu třetinu plochy souší. Pravděpodobně se jednalo o
nejvyšší zdvih hladiny světového oceánu v celém fanerozoiku. Významné
paleogeografické změny nastaly zejména po rozsáhlé cenomanské
transgresi, kdy globální vzestup mořské hladiny vyvolal značné zaplavení
níže položených částí pevnin a měl dalekosáhlé důsledky pro vývoj organického
světa. Koncem křídy naopak došlo k silnému poklesu mořské hladiny.
|
|
|
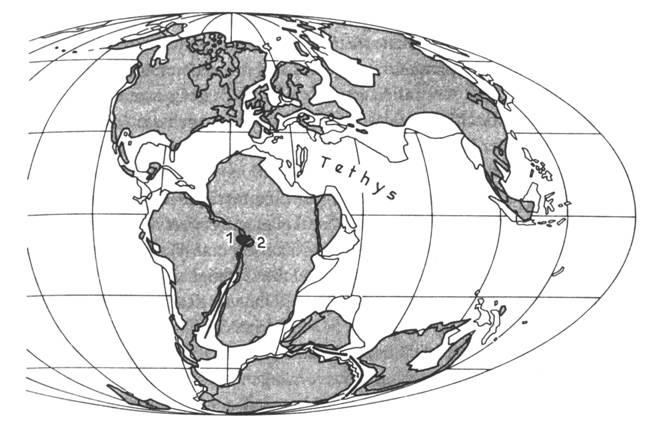
|
Obr. 8.3.1 Uspořádání kontinentů ve spodní
křídě (130 Ma). Pánve se sladkovodní spodnokřídovou
sedimentací: 1 – pánev Reconcavo,
2 – Gabunská pánev. |
Obr. 8.3.2 Uspořádání kontinentů ve svrchní
křídě (85 Ma). |
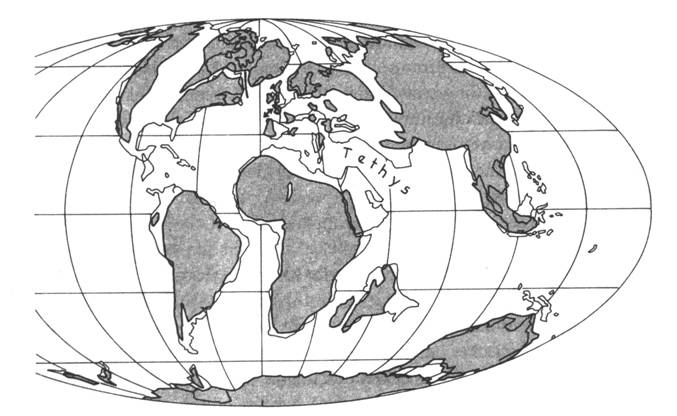
|
|
|
|
Obr. 8.3.3 Sediment svrchní křídy,
příbojová facie Kutná Hora, skalní město, Adršpachy. |
|


V oblasti jižní Evropy i
na jiných místech Země docházelo během křídy k významným horotvorným
pochodům, které měly počátky již v juře. Uzavřel se severní a
severozápadní okraj oceánu Tethys, proběhla tzv. kimmerská orogeneze. Posouvající se části zemské
kůry směrem na sever se zastavily o stará zpevněná jádra kontinentů (např.
Český masiv). Při subdukci – podsouvání oceánské kůry došlo k odtržení a
vyvrásnění lehčích sedimentů, především vápenců z mělčích částí oceánu Tethys. Jednalo se o alpinské
vrásnění. Alpinské vrásnění je považováno za nejvýznamnější tektonickou
událost geologické historie Země, která vyvrásnila evropský kontinent. Dalo za
vznik horským pásmům Alp, Karpat a dalších pohoří blízkého i vzdáleného
Východu. V karpatské soustavě se nacházejí i křídové uloženiny Beskyd a
severovýchodní Moravy. V severní Americe probíhala sevierská (spodní křída) a laramická
(svrchní křída) orogeneze, vzniklá důsledkem podsouvání pacifické oceánské
desky. V konsolidovaných oblastech se v křídě začínají projevovat
tzv. saxonské pohyby. V západních
částech Českého masivu se zakládají nové sedimentační pánve. Nejvýznamnější
z nich je česká křídová pánev.
Křídová fauna a flóra prošla
významnými změnami. V období svrchní křídy započal novověk rostlin, tzv. kenofytikum. Flóra ve spodní křídě byla obdobná
předešlé, jurské. Převažovaly rostliny nahosemenné, přesně cykasovité
a benetitové, dále byly hojné jehličnany a rostliny ginkgovité. Ve vzácnějších případech se vyskytovaly
rostliny kapraďosemenné. Na hranici spodní a svrchní křídy se již objevily
pokročilejší rostliny krytosemenné (Angiospermophyta). Během svrchní křídy dosáhly velkého
rozvoje rostliny krytosemenné obdobně jako v permu rostliny nahosemenné.
Ve svrchní křídě ovšem jen část přetrvala, především cykasovité
a v menší formě ginkgovité rostliny. Dnešní doby
se dožily jediné jehličnany. Vzniku krytosemenných rostlin napomohlo alpinské
vrásnění, a to změnou životních podmínek. Na konci spodní křídy mají nad
krytosemennými rostlinami převahu rostliny nahosemenné, flóra se podobá svrchní
juře. Na hranici spodní křídy a svrchní křídy (v cenomanu)
počet krytosemenných rostlin narůstá, a dokonce začínají nad nahosemennými
převažovat. Starší květenu zahubily pravděpodobně rozsáhlé mořské záplavy.
Stará květena nebyla schopná tak rychlého šíření na nových vynořujících se
pevninách, jako květena nová (rostliny krytosemenné). Zástupci křídových
krytosemenných rostlin představovali tropické nebo subtropické typy myrtovité,
magnólie, sasafrasy, liliovníky atd. Vyšší zeměpisné šířky obývala spíše flóra
mírnějšího pásma, jako jsou duby, buky, vrby, břízy a z nahosemenných
rostlin převládaly jehličnany v zastoupení araukirií,
sekvojů a borovic.
Nadměrný počet krytosemenných rostlin měl za
následek vymření býložravým dinosaurů. Úbytek těchto býložravých dinosaurů
zasáhl do potravního řetězce, tím pádem zastihl i nové predátory. Celkově během křídy dinosauři vymírali a
konce křídy se dožil jen zlomek. Příčiny je možné hledat v klimatických
změnách. Výjimku tvořily opeřené druhy dinosaurů současně s ptáky, kdy
jejich vznik souvisel s bohatým jídelníčkem v podobě hmyzu. Hmyzu se
dařilo díky rozvoji kvetoucích rostlin. Ptáci v období křídy vytěsnili ze
vzduchu létající plazy pterosaury.
Značné změny nastaly v mořském prostředí. Pokračoval rozvoj
prvoků, mezi které patří horninotvorné i stratigraficky významné skupiny dírkovců a bičíkovců. V teplých
mořích se dařilo živočišným houbám a
šestičetným korálům. Ramenonožci ustupovali. Rozvoj poznamenal
v křídových mořích mlži, kteří
se přisedlým způsobem života drželi hlavně na okrajích moře Tethys.
Typickým zástupcem mlžů byl rod Inoceramus. Hojně rozšíření byli měkkýši (především ústřice
a gastropodi), břichonožci,
spongie, ostnokožci (ježovky, hvězdice a hadice)
i korýši. Stále ještě důležitými
obyvateli křídových moří byli hlavonožci čtyřžábří (Ammonoidea) i dvoužábří
(Dibranchia),
avšak už i ti pozvolna mizeli. Někteří byli schopni se adaptovat na život při
mořském dně, kde vytvářeli rozvinuté stáčené i nestáčené schránky. Rod Parachydiscus měl
stočenou schránku, která mohla dosahovat velikosti až 2,5 m.
|
|
|
Obr. 8.3.4 Amoniti spodní křídy vnějších
Západních Karpat. |

|
|
|
|
Obr. 8.3.5 Crioceratites sp., na obvodu schránky jsou patrny ostny,sponí křída, Ladce, Slovensko. |
Obr. 8.3.6 Dufrenoya dufrenoyi., apt, Francie. |


|
|
|
|
Obr. 8.3.7 Duvalia sp., spodní křída, Slovensko. |
Obr. 8.3.8 Hibolites sp., barrem, Slovensko. |


|
|
|
|
Obr. 8.3.9 Inoceramus sp., Štíty, svrchní křída. |
Obr. 8.3.10 Micraster sp., spodní křída. |


|
|
|
|
Obr. 8.3.11 Protocallianassa antiqua, svrchní křída, Benátky u Litomyšle. |
Obr. 8.3.12 Žraločí zub, křída, Kolín. |


V mořích pokračoval rozvoj
žraloků, z tehdejších šestnácti
druhů se současnosti dožilo dvanáct. Rozvoj se týkal dále rejnoků i mořských
ryb. Hojnost ryb lákala do moří i četné plazy, kteří se přizpůsobovali životu
v moři. K nebezpečným dravcům křídových moří patří velcí plazi mosasaridi, kteří ovládli celou Zemi. Mosasauridi
byli vzhledově velmi podobní dnešním varanům, s tím rozdílem, že místo
končetin měli ploutve a tělo pokryto šupinami. Evropský rod Mosasaurus představoval ještěra
velikého až 12 m.
Ptáci se během křídy rychle
vyvíjeli, jejich tělo dosáhlo moderní podoby. V období spodní křídy se
vyvinuly první skupiny vačnatých a placentálních savců, kteří již nahradili
některé primitivní skupiny jurských savců. Vačnatci (Marsupialia) vzhledově
připomínali dnešní vačice. Žili na jižní polokouli, kdy byla propojena Jižní
Amerika, Afrika, Antarktida a Austrálie. Po rozdělení těchto kontinentů, které
proběhlo ve svrchní křídě, se již vyvíjeli samostatně. V krátkém období
eocén – miocén pronikli vačnatci i do Evropy. Placentálové
(Placentalia)
představují skupinu fosilních i recentních savců, kteří dokázali vytvářet
placentu v průběhu vývoje mláďat v těle matky. Tento orgán propojuje
zárodek se stěnou matčiny dělohy a dodává embryu živiny. Narozená mláďata byla
mnohem lépe vyvinuta jak mláďata vačnatců. Placentálové
vytvořili značné množství skupin, které dělíme do 29 řádů. Nejstarší skupinou
jsou hmyzožravci (Incestivora).
Pevniny prozatím ovládali
Dinosauři. Na africkém kontinentu již z jury vládl býložravý druh Brachiosaurus brancai, který
dosahoval délky až 22 m a výšky 12 m. Příbuzným rodem byl Brontosaurus a Diplodocus. Evropské dinosaury představují nejznámější iguanodoni. V severní
Americe v oblastech hojné vegetace žili neobvyklí rohatí dinosauři.
K nejznámějšímu rodu patří Triceratops. Další
méně známé rohaté dinosaury zastupují rody
Pentaceratos, Monoclonius a
Styracosaurus. Předchůdci rohatých dinosaurů byli objeveni v Mongolsku
v poušti Gobi. Nalezen byl přímo rod Protoceratopse, po kterém byly
nalezeny celé kostry a dokonce vejce, v nichž byla objevena embrya. Tento
objev dokazuje, že se dinosauři množili z vajec, která kladli do teplého
prohřátého písku. V Americe také žil ve svrchní křídě i podivuhodný
býložravý rod Trachodon
s tlamou ve tvaru kachního zobáku. Společně
s těmito dinosaury žili předchůdci dnešních pštrosů, dinosauři rodu Ornithoimus,
jejichž pozůstatky byly objeveny v také Gobi.
Masožraví dinosauři představují
nejmohutnější dravce, kteří kdy vůbec existovali na planetě Zemi. Největším a
zároveň nejznámějším představitelem masožravého dinosaura byl Tyrannosaurus.
Tento až 5 m vysoký dravý dinosaurus měl zakrnělé přední končetiny a zadní
končetiny tvořené třemi prsty. Tyrannosaurus se
živil masem jiných ještěrů. Příbuzný,
menším rodem byl Gorgosauru a
Megalosaurus. Velikost některých dinosaurů můžeme
odhadovat pouze z velikosti jejich stop, hojně jich bylo objevených ve
svrchní křídě státu Utah a Colorado.
Hranice
křídy/terciéru se vyznačuje velkým vymíráním, které postihlo
dinosaury (výjimkou větších opeřených typů), mořské ještěry, hlavonožce
(belemnity a větší část amonitů), mlže (rudisty a inoceramy). Hranice křídy/terciér nebyla na našem území
zachována. Existují dvě vysvětlení tohoto velikého vymírání. První teorie
připisuje vymírání na konci křídy pádu meteoritu v okolí Yucatánu a mexického zálivu (kráter Chicxulub).
Druhá teorie se spojuje s intenzivní vulkanickou činností v Indii,
pravděpodobně vyvolaná pádem dalšího velkého meteoritu (kráter Shiva). Nejznačnější změny v biotě jsou zřetelné na
západní polokouli. Podle nejnovějších interpretací pád meteoritu v oblasti
dnešního Mexického zálivu (Chicxlub) měl vliv jen na
dílčí biotu (hlavně Severní a Střední Ameriku, kdy důkazy tvoří sedimenty
tsunami), následovala intenzivní vulkanická činnost v Indii (Dekkan), která razantně ovlivnila klima a tudíž i biotu.