9. Kenozoikum
Poslední období vývoje naší planety
Země je označováno jako kenozoikum, anebo novověk Země. Éra trvající zhruba 65
miliónů let je obzvlášť významná především pro nás. Během kenozoika se vyvinula
skupina primátů (nehetnatců), ze které v poslední řadě vznikl i člověk.
Kenozoikum je charakterizované dovršením a ukončením alpínského vrásnění, které
zformovalo mladé pásemné pohoří a zemský povrch tak dosáhl dnešní podoby.
Eratém kenozoikum se dělí na tři útvary
paleogén, neogén a kvartér. Útvar paleogén je tvořen odděleními
označovanými jako paleocén, eocén a oligocén. Neogén se dělí na dvě oddělení, a
to miocén a pliocén. Poslední útvar kvartér tvoří oddělení pleistocén a
holocén. Mezinárodní dělení terciéru vychází z typických oblastí
v západní v Evropě (paleogén) a mediteránní oblasti (neogén). Za
třetihory (terciér) se považuje paleogén a neogén, a za čtvrtohory kvartér. Jako terciér „Montes
tertiarii“ označil Arduin v
r. 1760 útvar sypkých, nezpevněných sedimentů v podhůří italských Alp. Název kvartér
pochází z členění francouzského geologa J. Desnoyerse
z roku 1829, který po priméru, sekundéru
a terciéru označil nejmladší období zemské historie jako kvartér.
|
|
|
Obr. 9.1 Rozdělení Kenozoika. |

9.1 Paleogén (65,5±0,3 až 23,03±0,05 miliónů
let)
Paleogén započal globální krizí na hranici mezozoika a kenozoika, kdy je jeho spodní hranice daná pátým největším vymíráním v historii Světa. Stratotyp této hranice leží v El-Kef v Tunisu. Biostratigraficky leží tato hranice těsně při bázi zóny rozsahu foraminifery Parvularugoglobigerina eugubina, která představuje nástup terciérních planktonických dírkovců. Útvar paleogén se rozděluje na oddělení paleocén, eocén a oligocén.
Rozložení kontinentů se v paleogénu blíží dnešnímu stavu. Oproti křídě se rozšiřuje prostor Atlantického oceánu. Na jižní polokouli se definitivně oddělila Austrálie od Antarktidy. Tento proces umožnil chladné proudění kolem Antarktidy, což přispělo od eocénu k jejímu zalednění. Pokračuje pohyb indické desky k severu, která se v eocénu sráží s deskou asijskou.
|
|
|
Obr. 9.1.1 Uspořádání kontinentů ve středním eocénu (45 Ma). |
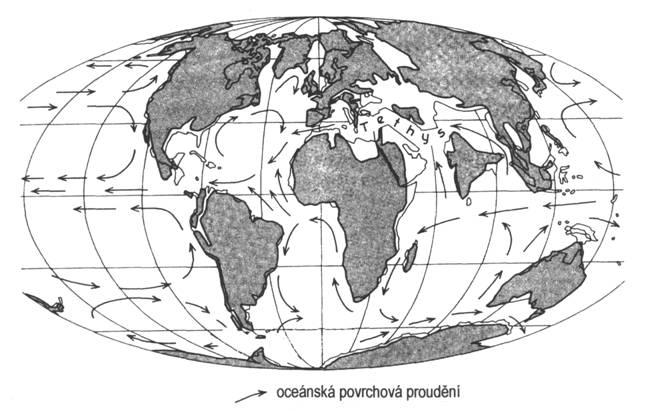
Alpinské vrásnění probíhalo od hranice křídy/terciér až do počátku eocénu prostřednictvím laramické fáze především v oblasti Severní Ameriky, také v oblasti Alp a Karpat. V blízkosti hranice eocén/oligocén se projevovala pyrenejská fáze, která působila hlavně v oblasti dnešní jižní Evropy. Konce paleogénu působila další sávská fáze, která tvořila Karpaty. Tyto výrazné pohyby zapříčinily rozpad severní části Tethydy. V paleogénu také proběhlo oddělení Jižní Ameriky od Afriky. Dále došlo k přesunu indického subkontinentu z jižní polokoule na severní, a následnému vyvrásnění Himalájí. Alpínské vrásnění ve východních oblastech Asie pokračuje i nadále.
Vývoj paleogénu v jižnějších částech Evropy ovlivnilo stěhování africké desky k severu. Toto tektonicky neklidné období doprovázejí další fáze alpinského vrásnění, které se zahrnují pod název neoalpinské vrásnění. V paleogénu k nim patří pyrenejská a sávská fáze. Pokračuje zvedání centrálního alpinského pásemného pohoří od Pyrenejí přes Alpy a Karpaty daleko na východ. Po obou jeho stranách v předpolí probíhá intenzivní flyšová sedimentace. Maximálního rozsahu mořské sedimentace bylo dosaženo v eocénu.
|
|
|
|
Obr. 9.1.2 Příklady flyšové sedimentace ve vnějších Západních Karpatech. |
|


V souvislosti s ukončením flyšové sedimentace v eocénu až oligocénu je mořská sedimentace vytlačovaná dále do předpolí, kde mohou vznikat nové sedimentační prostory (např. ve Vnějších Karpatech). V závěru paleogénu se západní část tethydního sedimentačního prostoru rozděluje na dvě samostatné větve – na jižní, vázanou na oblast dnešního Středozemního moře (tethydní větev), a na větev severní (paratethydní) v západním předpolí Alp, pokračující až do kaspické oblasti.
V konsolidovaných částech Evropy pokračují pohyby na starých i nových zlomových liniích, které se označují jako saxonské pohyby. Vznikají při nich též příkopové propadliny označované jako prolomy (rifty), např. rýnský, ruhrský, limagneský a ohárecký (též oherský) prolom. Jsou zaplňovány říčními a jezerními, místy i mořskými uloženinami. Na příhodných místech v nich vznikají hnědouhelné sloje. Do této kategorie lze přiřadit též vznik pohřbených praúdolí na jv. Moravě, označovaných jako vranovická a nesvačilská deprese.
V počátku paleogénu bylo podnebí velmi teplé, obdobné předešlému křídovému. Po prvním ochlazení proběhlo mírné oteplení. Na hranici paleocénu a eocénu bylo podnebí nejteplejší z celého kenozoika. Od toho oteplení docházelo již jen k postupnému ochlazení, které pokračuje do dnešní doby. Na konci eocénu došlo k oddělení Antarktidy a Austrálie, čímž vznikl cirkumantarktický mořský proud, který společně s poklesem CO2 v atmosféře zapříčinil celosvětové ochlazování. Během oligocénu se utvořil na Antarktidě ledovcový pokryv obestoupen tundrou. Na severní polokouli se zalednění prozatím nedostalo. V důsledku těchto změn vnikla výrazná roční období. Začala se šířit mírná pásma.
Flóra paleogénu byla již velice blízká té dnešní. Rozvoje dosáhly především dvě skupiny, a to moderní typy jehličnanů a krytosemenné rostliny. Jehličnany patřily k hlavním uhlotvorným dřevinám hnědouhelných močálů. Vegetace se dělila na dva typy: arktoterciérní opadavá převládající v oblastech Grónska, Kanady, Kamčatky a Špicberků, a druhou skupinu představují starostředozemní vždyzelená (Gelinden). V období eocénu pokračovala boreotropická (tropicko-subtropická) vegetace směrem na sever. Mírné ochlazení v oligocénu bylo propojeno s nástupem opadavých dřevin všech skupin, včetně bylin. Rostliny se do Evropy dostaly z Asie přes turgajskou šíji. V paleogénu v oblastech střední a severní Evropy se dařilo magnóliím, vavřínům, skořicovníkům, kafrovníkům, dubům, kaštanům, platanům, fíkovníkům, palmám atd. Vlhké prostředí bylo ideální pro mohutné bahenní tisovce (Taxodium) a naopak suchá místa pro obrovské sekvoje. Kmeny těchto rostlin tvoří třetihorní hnědé uhlí. V oblastech s mírnějším podnebím rostly jehličnaté stromy typu cedr, borovice a cypřiše. V pobaltských oblastech rostla významná borovice Pienus succinifera, jejiž zkamenělou pryskyřice nazývanou jantar, lze dnes nalézt na pobřežích Baltského moře. Jantary v sobě často ukrývají celé květy, komáry, mravence, brouky či pavouky.
|
|
|
Obr. 9.1.3 Pinus oviformis, Košťany (sbírka VŠB-TU Ostrava). |

Podstatné paleogeografické změny probíhající koncem křídy a začátkem paleogénu měly za následek i změny v životě různých živočichů. Vymírání postihlo mořské planktonní mikroorganismy s vápnitou schránkou, kam patří planktonní foraminifery a vápnitý nanoplakton. Následovala diverzifikace, která dosáhla maxima v období eocénu, avšak hned v oligocénu vlivem ochlazení diverzita mořského planktonu zase poklesla. Planktonní mikroorganismy mají důležitý stratigrafický význam a slouží k členění paleogenních mořských sedimentů, především se jedná o planktonní dírkovce rodu Subotina, Morozevella a Acarinina, vápnitý nanoplankton rodů Chiasmolithus, Discoaster, Sphenolithus a Reticulofenestra. Dále jsou stratigraficky významní mřížovci, rozsivky a dinoflageláta. Typickým zástupcem paleogénu byli velké druhy jednobuněčných prvoků - dírkovců rod Nummulites. Tento rod dírkovce byl hojně rozšířen v teplých a mělkých mořích po celém světě. Můžeme se s ním setkat např. ve vápencích, které tvoří pyramidy v Gíze.
|
|
|
|
Obr. 9.1.4 Numulitový vápenec, Rožumberok Slovensko; Tepe Kermen, Krym. |
|


|
|
|
Obr. 9.1.5 Clypeaster sp., paleogén. |

Na hranici křída/paleogén proběhl zvrat u suchozemské fauny. Období plazů bylo ukončeno a nastala doba hmyzu a savců. V období třetihor močály a jezera obývali v hojném počtu obojživelníci. V počátcích třetihor, kdy zavládnul velký rozvoj krytosemenných rostlin současně i rozvoj hmyzu, se také výrazně dařilo ptactvu. Dostatek potravy (nespočet hmyzu, semen a plodů) a hlavně žádná konkurence vytvořila ptákům dokonalé podmínky pro rozvoj. Byly zastoupeny již všechny současné řády. Existovaly i obrovské dravé nelétavé formy (Gastornis).
|
|
|
Obr. 9.1.6 Mene rhomba, eocén, Itálie (www.fossilmuseum.net). |

Souše rychle obývali savci. Vývoj savců na jednotlivých kontinentech nebyl jednotný, zaleželo na spojení pevninských mostů mezi kontinenty dovolující migraci. Z hlediska paleontologie se jedná o nejzajímavější kapitolu historie Země. Savci přicházející během třetihor byli velice přizpůsobiví, a protože neměli konkurenci a dokázali se rychle přizpůsobovat nejrůznějšímu prostředí, došlo u nich na počátku třetihor k obrovskému rozvoji. K rychlému rozvoji napomohl i rychlý rozvoj kvetoucích krytosemenných rostlin, na jejichž existenci byli některé skupiny savců závislé např. býložravé opice. Na konci spodní křídy ze starobylých všesavců (Pantotheria) se vyvinuli vačnatci a hmyzožravci. Žijící zástupci prastarých hmyzožravců jsou např. tanovití (Tupaiidea) žijící dodnes v Asii, ježci (čeleď Erinaceidae) a dále např. netopýři (Chiroptera).
Hlodavci se objevili na konci paleocénu, už v eocénu byli běžní v Americe i Evropě. Hlodavci bezesporu patří mezi nejúspěšnější a nejčetnější skupinu ze všech žijících savců. Dokáží se přizpůsobit všelijakému životnímu prostředí. Mezi vyhynulé skupiny paleogénu patří kreodonti (Hyaenodonta), kteří se podobali kočkám a psům a udrželi se až do oligocénu a prakopytníci (Protoungulata), kteří měli podobné znaky hmyzožravců, kopytníků a z menší části i šelem. K prakopytníkům patříla vymřelá pětiprstá skupina Amblypoda žijící v Asii i Severní Americe. Od eocénu se od nich odvozuje skupina lichokopytníků, kam patří tapíři (Lophiodon), pranosauři (titanotheria) a nejstarší zástupci koní. V Asii během oligocénu žil jeden z největších bezrohých příbuzných nosorožců, rod Baluchitherium. V té době to byl největší suchozemský savec, který měl protažený krk a přední nohy přizpůsobené ke spasení listí a v ramenou mohl dosahovat až 6 m. Vývoj koní započal v eocénu, kdy na počátku vývojové řady byl rod Eohippus, který dosahoval velikosti dnešního menšího psa a měl přední končetiny pětiprsté a zadní tříprsté. V Evropě se z něj vyvinul rod Palaeotherium, který ovšem dosahoval velikosti větší jak dnešní koně. Tato linie koní vymřela v Evropě ve starších třetihorách. Rod Eohippus nahradily v oligocénu rody Orohippus, Epihippus a Mesohippus. Počátkem eocénu došlo k oddělení sudokopytníků. Sudokopytníci (Artiodactyla) tvoří druhou obsáhlou skupiny kopytníků, která se dělí do tří podřádů: Suiformes (prasata a hroši), Tylopoda (velbloudovití) a Ruminantia (přežvýkaví, kam náleží jeleni, antilopy, žirafy, tuři, ovce a kozy).
Významná byla migrace z Asie do Evropy, která byla ovlivněná ochlazením na hranici eocén a oligocén a je označována, jako Grande Coupure. Objevují se pekariovití (Oreodon), prasovití (Suidae), moderní čeledi pravých šelem (Carnivora) i první předchůdci lidoopů Propliopithecus (oligocén). Savci během paleogénu také ovládli vzduch i vodu.
Ve svrchních třetihorách byli běžní chobotnatci (Proboscidea), mezi které patří sloni, mastodonti a mamuti. Chobotnatci vzniklí zřejmě v Africe, později putovali do Eurasie, Severní Ameriky a následně do Jižní Ameriky. Skupina chobotnatců byla velmi rozšířená a z hlediska paleontologie velmi významná, protože po sobě zanechala značné fosilní pozůstatky. Dnešní doby se dožili pouze dva zástupci, a to slon indický a slon africký. Nejstarším zástupcem mastodontů byl rod Palaeomastodon a Phiomia z oligocénu. V sedimentech usazených na hranici oligocén/miocén se po celé Euroasii, Africe a Americe objevuje dobře známý rod Trilophodon. Tento mastodont byl předchůdcem mnoha forem. Jejich nápadným znakem byly spodní řezáky, které představovaly nejrůznější tvary (např. lžícovité sloužící k rytí ve hlíně). Tyto rozdílné formy mastodontů vybudovaly dvě hlavní vývojové linie, které se lišily především tvarem stoliček. Ve svrchním miocénu u této linie proběhlo prodlužování horních řezáků tzv. klů i chobotu. Většina z nich však během pozdních třetihor vymřela. Poslední skupiny mastodontu žijících ve Střední a Severní Americe se dožily člověka a pouze jediná skupina přežila do současnosti a vytváří čeleď dnešních slonů (Elephantidae). K této čeledi patřil i nejbližší příbuzný dnešního slona indické, vyhynulý mamut Mammuthus.
9.
2 Neogén (23,03±0,05 až 2,588±0,005 miliónů let)
Útvar neogén se dělí na oddělení miocén a pliocén. Miocén obsahuje stupně aquitan, burdigal, langh, serraval, torton, messin. Pliocén má jen dva stupně, a to zancl a piacenz. Spodní hranice neogénu je vymezena na profilu Carossio-Lemme v severní Itálii výskytem foraminifery Paragloborotalia kugler. Svrchní hranice je nově od června roku 2009 definována na bázi stupně gel na profilu Monte San Nicola na Sicílii.
V evropském členění neogénu na stratigrafické stupně se v posledních padesáti letech uplatňuje dvojí dělení: stupně definované v mediteranní (tethydní) oblasti a stupně stanovené v centrální Paratethydě. Rozdílnost dělení se neprojevuje jen v odlišných názvech stupňů, ale rovněž v rozdílném pojetí spodní hranice neogénu a v nesouhlasném vedení hranice mezi miocénem a pliocénem. V nedávné době k dosavadnímu dělení pliocénu na dva stratigrafické stupně v mediteranní oblasti přibyl ještě nejmladší stupeň s názvem gelas (stratotyp na Monte St. Nicola u Gely na Sicílii).
Dvojí členění neogénu má značný dopad na stratigrafické členění neogénu v západokarpatské oblasti na našem území a ve vídeňské pánvi. V starších dobách byly v této oblasti aplikovány mediteranní (tethydní) stupně, posléze pak nové stupně paratethydní. Vede to k určitým obtížím a nesrovnalostem ve starší a mladší literatuře. Z tohoto pohledu bude na tomto místě vhodné poznamenat, že v naší starší literatuře se používalo označení burdigal ve smyslu paratethydního stupně eggenburg, později opuštěný termín helvet pro nynější ottnang a karpat a konečně označení torton pro stupeň baden.
V současné době je ovšem za mezinárodní standard považováno stupňové dělení neogénu stanovené v mediteranní oblasti. Paratethydní dělení na eger až roman jsou považovány za regionální stupně.
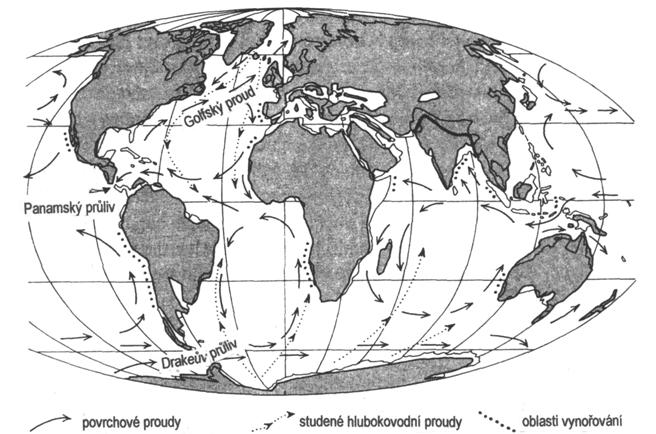
V období miocénu došlo k mírnému oteplení a vysušení podnebí. Svrchní miocén je označován za nejteplejší období za posledních 35 miliónů let. Ve svrchním miocénu byla Antarktida značně pokryta ledem, což způsobilo glacieustatický pokles hladiny moří, který byl zakončen na konci miocénu izolací a vysoušením Mediteránu (označováno jako messinská salinitní krize). Za ochlazení započaté ve svrchním miocénu pravděpodobně mohlo propojení Atlantiku s arktický mořem, které v tu dobu sloužilo jako globální tepelný výměník. Současně se zvýšila nadmořská výška kontinentů. Vyšší teploty ve středních zeměpisných šířkách ovlivňovaly značné rozšíření tropického a subtropického lesa.
Alpinsko-himalájská orogeneze nadále přetrvávala. Sávská fáze z oligocénu přetrvávala do miocénu, pak byla nahrazená fází štýrskou, která byla vystřídána fází attickou (ta sestavila kontinenty a moře téměř do takové podoby jak vypadají dnes). Alpinská orogeneze dosáhla maximu během rhodánské fáze, po ní už byla zjevná horotvorná činnost ukončena. Menší pohyby probíhají do dnes. Tektonické napětí vyvrásnilo mohutné horské pásma Kordiller v Severní Americe, And v Jižní Americe a Himalájí v Asii. Tyto tektonické procesy doprovázel vznik příkopových propadlin. Současně docházelo k posunu bývalých gondwanských kontinentů. Afrika společně s Arabským poloostrovem byla nasunuta na Evropu, čímž vytvořila přírodní hráz v místech dnešního Gibraltaru a ze Středozemního moře se tak stalo bezodtokové slané jezero s rozměrnými zásobami evaporitů. Přesun Indie na sever zapříčinil uzavření Tibetského moře a vyvrásnění Himalájí. Na konci miocénu se snížila mořská hladina, která způsobila propojení některých kontinentů pevninskými šíjemi. Nova Guinea s Austrálií zformovali jednotný kontinent. Asie se propojila s Aljaškou. Během pliocénu došlo ke spojení Severní a Jižní Ameriky, což výrazně ovlivnilo endemickou faunu a pravděpodobně tento pevninský most narušil atmosférickou cirkulaci a teplé proudy odklonil k severu. Mohl to být jeden z hlavních faktorů, které zřejmě vytvořil mohutné ledovce na severní polokouli, a také pravděpodobně ovlivnil střídání dob ledových a meziledových během pleistocénu.
Rozložení kontinentů a oceánů v neogénu je blízké dnešnímu stavu (obr. 16). Západní část Tethydy v evropském prostoru je od konce paleogénu rozdělena na dvě větve: na jižní – mediteranní a na větev severní, která se označuje novým názvem Paratethys. V jižní větvi neogénní sedimentace pokračuje v prostoru Středozemního moře. Severní větev má zpočátku souvislý transevropský průběh. Můžeme ji sledovat od západního předpolí Alp až do kaspické oblasti. Vzhledem k její délce a k některým specifikám v geologickém vývoji se rozlišuje západní, centrální a východní Paratethys.
|
|
|
Obr. 9.2.1 Evapority badenu (především halit); Bochnia Polsko. |

Významnou
událostí na konci miocénu, která ovlivnila paleogeografický vývoj především
v jižní Evropě, je tzv. messinská krize. V důsledku dočasné izolace
prostoru Středozemního moře ve stupni messin došlo
k jeho vysýchání. Na jeho dně se vylučovaly až přes
|
|
|
|
Obr. 9.2.2 Uspořádání kontinentů ve středním miocénu (12 Ma). |
|
|
|
|
|
Obr. 9.2.3 Paleogeografická rekonstrukce střední Evropy během spodního miocénu (s použitím mapy P. A. Zieglera). |
|
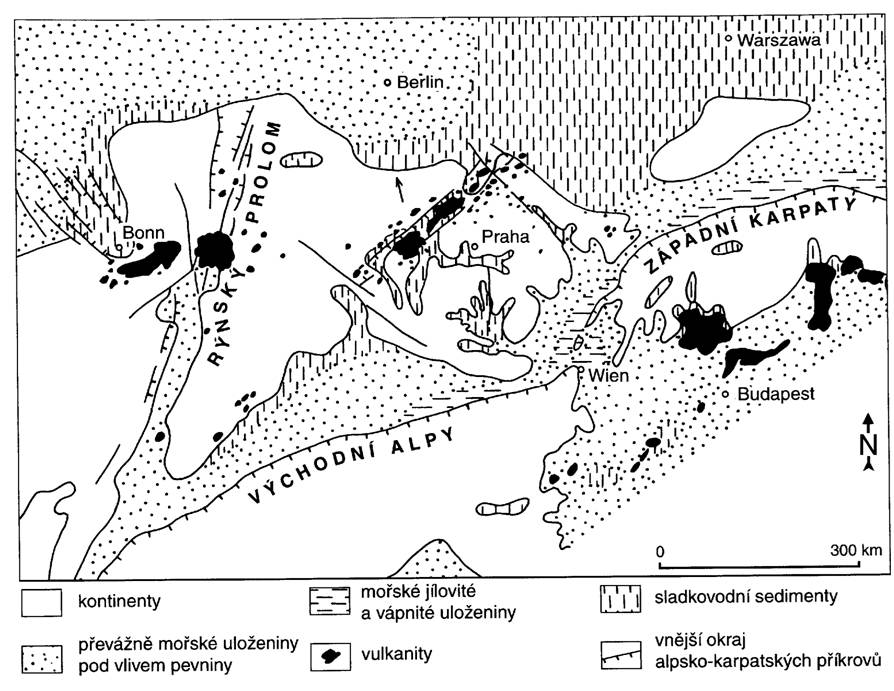
Ranější sedimentace v Paratethydě probíhala pod přímým vlivem dalších fází neoalpinského vrásnění. Na území České republiky se jako nejsilnější z nich uplatnila mladoštýrská fáze v badenu, která je v centrální Paratethydě (ve Vnějších Západních Karpatech) hlavní příkrovotvornou fází. Na neogénní uloženiny západokarpatské předhlubně byl nasunut komplex vněkarpatských (flyšových) příkrovů.
|
|
|
|
Obr. 9.2.4 Sedimenty eggenburgu vídeňské pánve. |
|


Zatímco v tethydní oblasti probíhaly další procesy neoalpinského vrásnění, v konsolidované části Evropy pokračovaly saxonské pohyby na zlomových liniích. Často byly doprovázeny uhlotvornou sedimentací a intenzivním suchozemským výlevným vulkanismem. Společným znakem vulkanitů je alkalický charakter a převaha bazických až ultrabazických hornin. Vulkanické projevy jsou často sledovatelné až do kvartéru.
|
|
|
|
Obr. 9.2.5 Mostecká uhelná pánev. |
|


|
|
|
Obr. 9.2.6 České středohoří. |

Během spodního miocénu se na naše území vrátily paleotropické prvky rostlinstva, např. zástupci čeledi Mastixiaceae, Fagaceae, Lauraceae a palmy. Následné ochlazení vystihují formace smíšeného mezofytního (vyžadující mírnou vlhkou půdu) lesa. V pliocénu se dařilo bylinné vegetaci, ovšem jen do té doby než nastoupila ledová období. Teplomilné prvky postupovaly k jihu a jihovýchodu (Itálie, Zakavkazí: buk, dub korkový, vavřínovité, palmy), ovšem v důsledku bariéry ve formě alpsko-karpatského horstva se stěhování jižním směrem nekonalo. Velká část terciérních reliktů se v Evropě vůbec nezachovala, pouze v Severní Americe a východní Asii. Neogenní rostliny slouží k paleoklimatické rekonstrukci.
|
|
|
|
Obr. 9.2.7 Taxodium dubium, spodní miocén (sbírka VŠB-TU Ostrava). |
|


Mořský plankton slouží jako nejlepší biostratigrafický indikátor pro terciérní sedimenty. Pomocí mořského planktonu (především podle planktonních dírkovců, vápnitého nanoplanktonu, rozsivek, mřížovců, dinoflagelát) je možné rozdělit mořské sedimenty paleocénu až holocénu na celosvětově platné zóny. Skupiny organismů s vápnitou schránkou se rozvíjely rychleji. Je zjištěno, že průměrná délka zóny vápnitých organismů je 1-2 milióny let, křemičitých organismů 3-4 milióny let. Původní mořské houby, které vyhledávaly mělká moře, již vymizely a udržely se jedině ty, které měly kostry z oxidu křemičitého a obývaly větší hloubky. Dále byli zastoupeni útesotvorní i solitérní koráli, mechovky, gastropodi, břichonožci a méně pak hlavonožci a ježovky. Měkkýše zastupují hlavně rody mlžů Pecten, Congeri a Unio. Poměrně hojní byli i krabi, rody Coleoma, Balanus.
|
|
|
|
Obr. 9.2.8 Pecten sp., neogén. |
Obr. 9.2.9 Glycimeris sp., neogén. |


|
|
|
|
Obr. 9.2.10 Conus sp., Mikulov, miocén. |
Obr. 9.2.11 Melanopsis sp., Čejč, neogén. |


|
|
|
Obr. 9.2.12 Congeria sp. |

|
|
|
|
Obr. 9.2.13 Příklady neovulkanických projevů; Panská skála u Kamenického Šenova; lávový proud u obce Mezina. |
|


Ve Francii, Řecku, Pákistánu, Indii, Číně, na Kavkazu i v Keni byl dokázán život několika rodů čeledi Pongidea (lidoopi). Do této čeledi spadají dnešní šimpanzi, gorily i orangutani. Nejstarší představitelé této biologicky vyspělé čeledi pocházejí z počátků miocénu. Na hranici středního a svrchního miocénu se objevuje v oblasti Eurassie Sivapithecus, ten představuje zhruba 1,5 vysokého lidoopa obývající koruny stromů. V Pákistánu, Indii a v severní Číně se vyskytoval ramapiték (Ramapithecus) menší jak sivapiték, s hlavním rozdílem, že život trávil převážně na zemi. Vzpřímená chůze byla u něj běžná, mohl tak viděl přes vysokou trávu tehdejší stepi a přecházet řeky. Nepotřebné ruce tak mohl používat k uchopení nástrojů. Jeho chrup už nebyl čistě opičí, měl už znaky chrupu lidského. Podle nových studií ramapitéci a sivapitéci tvořili vývojovou linii, která vede k dnešním orangutanům. V pliocénu se objevuje na území dnešní Číny a Pákistánu zdatný lidoop Gigantopithecus velikostí dnešní gorily a váhy až 300 kg. Gigantopithecus patřil mezi býložravce. Domněnkou je, že se jedná o potomka bájného Yettiho (známého také pod jménem sněžný muž), který se,, údajně“ objevuje v Himalájí.
Velké množství řádů savců vymírá během konce miocénu. Hlavní příčinou je velký konkurenční tlak, který nastal pevninským propojením mezi Severní Amerikou a Asií, mezi Afrikou a Eurasií, mezi Severní a Jižní Amerikou. Zvířatům bylo umožněno se pohybovat mezi kontinenty, a tak musely o životní prostor bojovat s domácími druhy. Negativně také přispívalo zhoršující se podnebí. Ze skupin savců tyto konkurenční boje nepřežila řady kopytníků (např. řády Astrapotheria, Cainotherioidea atd.).
9.3
Kvartér (2,588±0,005 miliónů let až současnost)
Kvartér představuje nejmladší a současně nejkratší období v historii Země. Kvartér zaujímá významné postavení, a to především ze dvou důvodů. Prvním důvodem je rychlý pokles teploty, který má za následek značné rozšíření ledovců. Druhým důvodem je vývoj moderního člověka, který toto období značně ovlivňuje. Kvartér lze rozdělit na dvě oddělení: starší pleistocén (doba ledová) a mladší holocén (doba poledová či postglaciál). Spodní hranice kvartéru je stanovena na bázi stupně gela na profilu Monte San Nicola na Sicílii. Hranice pleistocénu a holocénu nemá stratotyp, ale je dána na základě změn v grónském ledovci na úrovni 11 700 let př. n. l., kdy zavládlo teplejší klima. Přijetí této hranice je v rukách kvarterní stratigrafické komise.
Období kvartéru se vyznačuje střídáním glaciálů s interglaciály. Glaciály neboli doba ledová označuje chladné a suché období, během kterého jsou rozšířeny ledovce. Interglaciály neboli doba meziledová je teplejší období mezi dvěma glaciály. U kvarterních převážně terestrických sedimentů není známý úplný profil celým pleistocénem, a tak vzniklo množství lokálních, navzájem značně obtížně srovnatelných stratigrafických schémat. Jednotné a všeobecně platné stratigrafické schéma pleistocénu prozatím neexistuje. Kvartér je považován za období velmi dynamické, z důvodů cyklického střídání klimatu. Klimatické změny ovlivňují charakter sedimentů. V glaciálech se průměrná roční teplota pravděpodobně pohybovala kolem 0°C a šla zřejmě i níž, v interglaciálech ve stejné oblasti dosahovala 10-15° C. Dnešní průměrná roční teplota u nás dosahuje 8-9° C. Tyto klimatické výkyvy ovlivňovaly: posun klimatických pásem od severu k jihu a zpět spojený se změnami rozšíření ledovců, kolísání hladiny světového oceánu (glacieustáze) a střídavé zaplavování pobřežních oblastí, zatěžování a následné odlehčování pevnin ledovci, následná migrace fauny i flóry, střídání období rozrušování hornin (eroze a denudace) a hromadění sedimentů (akumulace). Nejmladší období kvartéru reprezentuje nástup současných společenstev a je možné při jeho studiu uplatnit princip aktualismu, tedy opírat se o poznatky současných probíhajících procesů na poměry v minulosti. Po ukončení nejmladšího zalednění započalo období dnešního postglaciálu tzv. holocénu ve kterém žijeme.
|
|
|
Obr. 9.3.1 Rozsah celosvětového zalednění v pleistocénu (http://jan.ucc.nau.edu/~rcb7/Pleistmoll.jpg). |
|
|
|
Obr. 9.3.2 Rozsah kontinentálního ledovce v Evropě v období před 20 tisíci lety (http://www.geocities.com/reginheim/europeiceage.gif). |
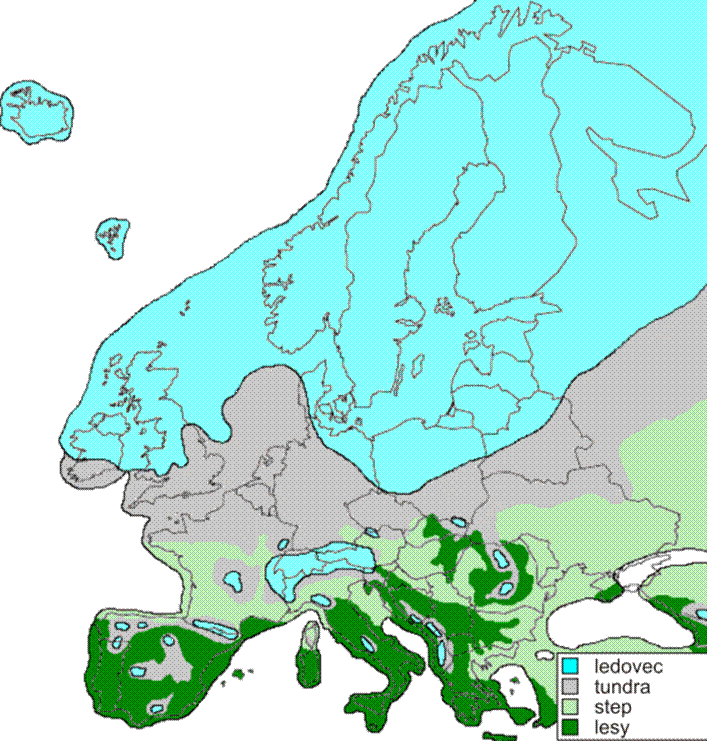
Ve střední Evropě se projevilo jak zalednění kontinentální (skandinávské), tak horské (alpinské). V alpinské oblasti se uvádějí 4 glaciály (gűnz až wűrm), v kontinentální obvykle jen tři hlavní (elster až weichsel). Glaciálu elster (též halštrov) kontinentálního zalednění odpovídá mindel; saale odpovídá riss a weichselu odpovídá wűrm. Mezi nimi se nacházejí interglaciály.
V ledových dobách se v předpolí severského ledovce rozprostírala ve velikosti 200-300 km tři vegetační pásma: tundra, step a tajga. Tundry převládaly v periglaciálních oblastech střední Evropy. Mezi typické rostliny tundry patří mechy, lišejníky (lišejník sobí), zakrslé polární vrby (Salix polaris), zakrslá bříza (Betula nana) a především typická plazivá růžovitá rostlina dryádka osmiplátečná (Dryas octopetala), podle které se tundrová flóra ledových dob nazývá jako flóra dryasová. Jižním směrem od polární tundry se rozkládalo pásmo tajgy s rostlinami jehličnatými a listnatými, především borovicemi, smrky, modříny a břízy.
V našich šířkách mělo rostlinstvo meziledových dob jiný charakter, buď vyhynulo, nebo se zachovalo pouze v jižní Evropě, dále také na Kavkaze či v jihovýchodní Asii. Patří sem např. leknínové rody Euryale a Brasenia, z šáchorovitých rod Dulichium, penišník Rhododendron ponticum, zimostráz Buxus sempervirens, ořešák Juglans regia, fíkovník Ficus carica atd. Některé rostliny např. kotvice vzplývavá (Trapa natans) a tis (Taxus baccata) hojně rostoucí v dobách meziledových, rostou dnes ve stejných oblastech zcela výjimečně. Nejjižněji se rozprostíraly oblasti smíšených doubrav s lískou, dubem, jilmem, javorem, lípou, bukem a jedlí. Pásma těchto rostlinných společenstev se posouvala od jihu k severu i naopak, na základě klimatických změn.
V interglaciálech, při nástupu vlhkého a teplého klimatu se dařilo lesnímu společenstvu. Ve spodním pleistocénu se udržely z terciéru rostlinné druhy: ořechovec Carya, jedlovec Tsiga, šácholan Magnolia, břestovec Celtis (dodnes žije ve Středozemí). Flóra středního a svrchního pleistocénu již má dnešní charakter.
|
Tabulka 9.3 Stratigrafické schéma kvartéru podle evropského dělení. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Pro období glaciálu je charakteristická megafauna, kterou zastupují živočichové, jak už plyne z názvu větších rozměrů. Mezi tyto skupiny živočichů patří např. chobotnatci: mamuti, mastodonti; lichokopytníci: nosorožci, Elasmotherium; sudokopytníci: Megaloceros, jihoameričtí edentati Megatherium, Glyptodon, Scelidotherium. Pleistocenní faunu zastupovali především savci - chobotnatci (Proboscidea). Největším vymřelým evropským slonem byl pětimetrový pleistocenní rod Palaeoloxodon, který obýval lesy v dobách meziledových. Blízkým příbuzným menšího vzrůstu byl rod Palaeoloxodon melitensis obývající Sicílii a Maltu. Ovšem nejznámějším pleistocenním představitelem byl jednoznačně mamut (Mammuthus primigenius) význačný představitel glaciální fauny. Tento mamut, jehož maximální rozvoj proběhl ve svrchním pleistocénu, vládl v tundře. Jeho pozůstatky byly objeveny na mnoha místech v trvale zmrzlé půdě (v permafrostu) na Sibiři, a jelikož byly nalezeny ve velmi dobře zachovalém stavu, bylo možné studovat i měkké části jeho těla. Tyhle nálezy také dokazují, že byl pokryt hustou hnědou srstí chránící před mrazem. Během nejmladšího pleistocénu byl značně rozšířen. Mamuti nežili samostatně, ale žili ve stádech. V době nejmladšího glaciálu se podařilo mamutovi migrovat skrz v té době vyschlou nebo zmrzlou Beringovu úžinu do Severní Ameriky, kde se mu dařilo a velmi se zde rozšířil. Ve starším pleistocénu obýval Severní Ameriku blízký druh Mammuthus imperator a dále také o něco menší kolumbijský slon Mammuthus columbi. Současně se v pleistocénu vyskytovali typičtí zástupci interglaciální doby, nosorožci Dicerorhinus etruscus a Dicerorhinus kirchbergensis, kteří se vyskytovali společně se slonem Paleoeloxodon antiquus. V dobách ledových vedle sebe žil mamut a nosorožec srstnatý Coelodonta antiquitatis. K velkým pleistocenním savcům patřili dále lichokopytníci, koňský rod Equus. Tento rod se vyvinul v Severní Americe, avšak rozšířil se do celé Asie i Evropy. Nejstarší zástupci rodu Equus v Evropě byli velice blízcí dnešní zebře, druh Equus stenonis žil na přelomu pliocénu a pleistocénu.
V teplejších obdobích kvartéru v Evropě žil hroch (Hippopotamus). Sudokopytníky dále zastupuje kvarterní jelen rodu Cervus. Jeho nejzajímavější zástupce byl obrovský jelen irský Megaceros giganteus s parohy, které měly v rozpětí až 3 metry. Pozůstatky tohoto jelena byly nalezeny i na našem území. V nejmladší době ledové byli běžní v Evropě sobi (Rangifer arcticus a Rangifer tarandus). Dále k sudokopytníkům náležel předchůdce dnešního skotu, a to pratur Bos primigenius. Běžný byl také zubr. V období glaciálu se vyskytoval pižmoň Ovibos moschatus a v pozdějších čtvrtohorách ve středozemní oblasti velbloudi.
V období pleistocénu byla Evropa domovem mnoha šelem, především medvědům. V předposlední době ledové se na našem území vyskytoval obrovský medvěd jeskynní Ursus spelaeus a jeskynní hyena Crocuta spelaea, kteří se udrželi do poslední doby ledové. Také pozůstatky šavlozubého tygra Machairoidus s výraznými špičáky byly objeveny na několika místech Evropy i u nás. Šavlozubý tygr je znám ze staršího pleistocénu. V Americe žil obdobný tygr rodu Smilodon. Během středního a svrchního pleistocénu žil na území střední a jižní Evropy jeskynní lev Panthera spelaea. O jeho životě dokládají kresby v jeskyních předvěkého člověka. Z dalších šelem se u nás žily v dobách ledových lišky, vlci, stepní tchoři, rosomáci, lumící (rod Myodes), syslové Citellus, bobři Castor fiber atd. V sedimentech dob ledových se objevují zbytky různých ptáků, např. kura rousného, kura alpského, některý druhy sov či pěnkav.
V kvartéru vygradoval vývojový proces, který započal u primitivních primátů a skončil u moderního člověka. Člověk vzešel z úzkonosých opic Starého světa, od kterých se koncem třetihor odpojila nová vývojová linie nadčeleď Hominoidea, kam patří lidoopi i lidé. Tato nadčeleď se dále dělí do čeledí: Oreopithecidae, Pliopithecidae, Pongidae a Hominidae. Čeleď Oreopithecidae a Pliopithecidae jsou čeledi vymřelé. Do čeledi Ponginae patří dnešní orangutani, šimpanzi a gorily. Mezi nejvýznamnější skupinu patří čeleď Hominidea (lidé), ke které patří člověk se svými předchůdci. Tato čeleď k rozvoji potřebovala pouhých 14 miliónů let a obsahuje v současnosti čtyři rody: Ramapithecus, Austrolopithecus, Paranthropus a Homo. Rod Ramapithecus, z Indie a Číny popisuje Ramapithecus punjabicus, a z afriky Ramapithecus wickeri. Tento rod představuje člověka zhruba 110 cm vysokého, pohybujícího se po dvou zadních nohách. Patří mezi nejstarší předchůdce člověka, který žil před 12 milióny lety.
Z východní a jižní Afriky pocházejí četné nálezy rodu Australopithecus africanu. Austrolopitékové byli vysocí zhruba 120 cm, pohybovali se po dvou končetinách a měli vzpřímenou postavu. Před cca 3,3 milióny lety obývali roviny či stepi a zřejmě byli schopni již ulovit větší zvířata, na která vyrazili v tlupách. Je pravděpodobné, že dokázali z pazourků vyrábět nástroje a dokázali používat oheň. Patřili již k vývojově pokročilejší skupině. Z Afriky jsou známé také nálezy rodu Paranthoropus, který má dva rody Paranthoropus robustus a Paranthoropus boisei. Oproti předešlému rodu australopitéků byl to rod vyšší a výrazně mohutnější, který obýval lesy a upřednostňoval rostlinnou stravu. Měl mohutnou lebku. Rod Paranthoropus žil v období mezi 2,7 -1,5 milióny let.
Rod Homo obsahuje všechny jak vyhynulé tak i žijící populace člověka. Řadí se sem Homo habilis, Homo erectus (poddruhy Homo erectus erectus, Homo erectus modjokertensis, Homo erectus pekinensis, Homo erectus lantianensis, Homo erectus heidelbergensis, Homo erectus palaeohungaricus, Homo erectus mauritanicus, Homo erectus leakeyi a dále Homo sapiens (poddruhy Homo sapiens steinheimensis, Homo sapiens palestinensis, Homo sapiens rhodesiensis, Homo sapiens soloensis, Homo sapiens neanderthalensis a Homo sapiens sapiens.
Nejstarším zástupcem rodu Homo je druh Homo habilis, který žil na africkém kontinentu zhruba před 2-1,6 milióny lety. Tento první pravý člověk dosahoval velikosti 130 cm a vážil přibližně 30-42 kg. Byl to člověk vzpřímené postavy pohybující se po dvou končetinách. Byl zručný, vyráběl a používal kamenné nástroje. Z Jávy jsou známy dva poddruhy rodu Homo erectus, a to primitivní forma Homo erectus modjokertensis a pokročilejší forma Homo erectus erectus. Podle tvaru nalezené stehenní kosti měl postavu vzpřímenou a tento člověk dosahoval výšky až 165-170 cm. V Číně byl objeven poddruh Homo erectus pekinensis a Homo erectus officinalis. Z Evropy pocházející svědectví o nejstarších zástupcích rodu Homo patří poddruhu Homo erectus heidelbergensis neboli člověku heidelberskému. Z Maďarska pochází mladší a pokročilejší forma Homo erectus palaeohungaricus. Z Ternifine a Severní Afriky mladší vývojový druh Homo erectus mauritanicus a ze severní Tanzanie Homo erectus leakeyi. Všechny poddruhy Homo erectus používaly kamenné nástroje, které u nich byly objevené, a u mnohých z nich byl prokázán dokonce kanibalismus.
Nejvýznamnějším druhem je Homo sapiens s plně vzpřímenou postavou a držením těla, které odpovídá dnešnímu člověku. Délka kostí jeho končetin odpovídá i těm dnešním. Představoval poslední vývojovou etapu člověka. Nejstarší představitel poddruhu Homo sapiens steinheimensis byl objeven v Německu a Anglii, a stáří se odhaduje na 250-400 tisíc let. Z Izraele, ze Zimbabwe, z ostrova Jávy známe poddruhy Homo sapiens palestinensis, Homo sapiens rhodesiensis a Homo sapiens soloensis. Nejmladším poddruhem je Homo sapiens sapiens, objevený zhruba před 40-30 tisíci lety, ke kterému patří dnešní člověk.
Z genetického hlediska dělíme kvartérní sedimenty do několika skupin. Vedle oblastí přímořských, kde se uplatňuje mořská sedimentace, převažuje na pevninách v zaledněných i v nezaledněných oblastech terestrická sedimentace se svými specifickými vlastnostmi.
Česká republika v pleistocénu ležela v dosahu obou typů zalednění, tj. mezi kontinentálním severoevropským zaledněním na severu a velehorským (alpinským) zaledněním na jihu. V tohoto hlediska na našem území rozlišujeme kvartérní sedimenty oblastí překrytých ledem, tj. oblastí glaciálních, oblasti příledovcové neboli periglaciální a oblasti nezaledněné neboli extraglaciální.
Severský neboli kontinentální ledovec zasáhl naše území dvakrát – v zalednění elsterském a salském. Glacigenní sedimenty v České republice nalézáme ve Šluknovském a Frýdlantském výběžku, na severní Moravě a ve Slezsku. K ledovcovým uloženinám patří především tily (s eratickými horninami), glacifluviální a glacilakustrinní sedimenty.
|
|
|
|
Obr. 9.3.3 Komplex glacifluviálních sedimentů. Sekundárně porušen kryoturbací. |
|

|
|
|
|
Obr. 9.3.4 Till (flow till), písčitý štěrk s bloky nordik. |
Obr. 9.3.5 Proglaciální sedimenty, sedimenty výplavové plošiny (sandru). |


{kind=link}
{kind=link}
Horské zalednění se mohutně projevilo v Alpách, odkud dosáhlo až na Šumavu. Pozůstatky horského zalednění u nás nacházíme ještě v Krkonoších, v Hrubém Jeseníku, Králickém Sněžníku a v Moravskoslezských Beskydech.
Značná část plochy našeho území v pleistocénu patřila periglaciální zóně. V glaciálech se zde výrazně uplatňovaly exogenní procesy mechanického zvětrávání. Ve vrcholových oblastech našich hor v pevných horninách vznikaly mrazové sruby s kryoplanačními terasami, které přecházejí na svazích v kamenná moře a suti.
V akumulačních oblastech uvedené zóny a v extraglaciálních oblastech k dominantním uloženinám patří říční terasy, spraše a váté písky. Významné jsou též fosilní půdy.